皮膚屏障 (Skin Barrier)
核心概念
- 皮膚最重要功能之一是在生物體與外在環境間形成屏障,保護身體免於乾燥 (desiccation)、物理應力、感染、過熱/散熱與紫外線 (UV) 傷害。
- 屏障主要由表皮 (epidermis)(角質化複層上皮)承擔,關鍵為角質層 (stratum corneum) 與緊密連接 (tight junctions)。
- 角質層=空氣–液體界面屏障,雙向防護:防水分散失(由內向外 inside–outside)+防有害物質進入(由外向內 outside–inside)。
- 緊密連接=液體–液體界面屏障,封閉顆粒層第二層 (SG2) 細胞間隙,限制分子經細胞旁路徑 (paracellular pathway) 移動。
表皮與角質層基本結構
- 表皮活性細胞層由內而外:基底層 (stratum basale)、棘層 (stratum spinosum)、顆粒層 (stratum granulosum)。顆粒層至少三層,由外而內為 SG1、SG2、SG3;緊密連接封閉 SG2 層。
- 表皮細胞僅在基底層增生,向上分化、於顆粒層扁平、SG2 形成緊密連接、SG1 失去緊密連接並終末分化為角質細胞 (corneocytes),最後以鱗屑脫落 (脫屑 desquamation)。持續更新使附著的外來物質隨脫屑被丟棄。
- 人類角質層厚約 10 至 20 µm,含約 10 至 25 層角質化細胞。
- 角質細胞靠角質橋粒 (corneodesmosomes) 與細胞間脂質層板的黏附力相互黏附。
- 常規 H&E 石蠟切片中角質層呈「籃網狀結構 (basket-weave structure)」,此為二甲苯去蠟萃取脂質造成的人為假象 (artifact),並非真實多孔結構。
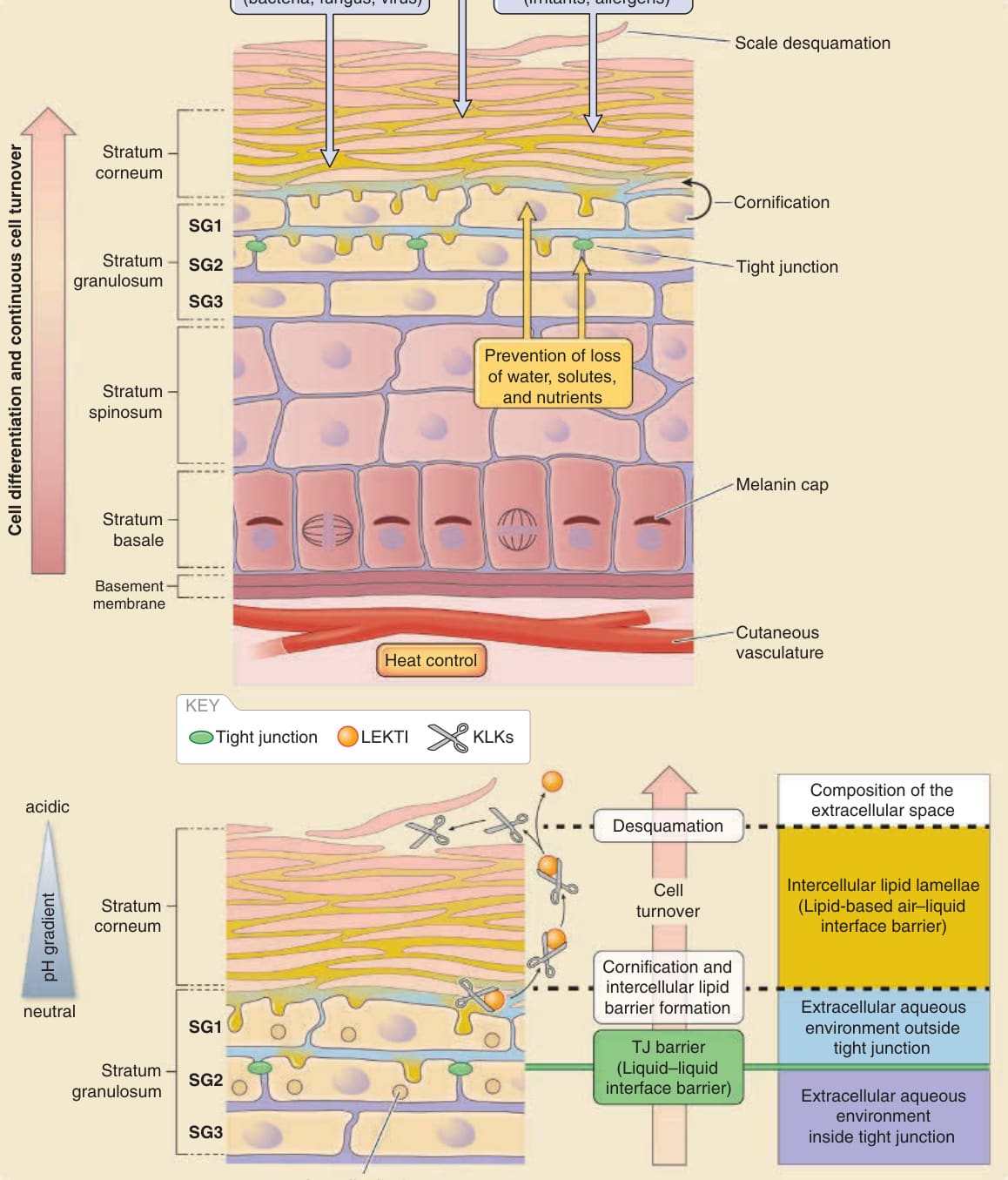
圖 14-2:表皮基本結構與主要表皮屏障
角質化細胞包膜與角質細胞脂質包膜
- 角質化時於質膜下方形成約 10 nm 厚、高度交聯不溶蛋白的角質化細胞包膜 (cornified cell envelope);質膜脂質雙層被約 5 nm 厚醯基神經醯胺 (acylceramides) 取代,即角質細胞脂質包膜 (corneocyte lipid envelope)。
- 兜膜蛋白 (envoplakin)、外周斑蛋白 (periplakin)、兜甲蛋白 (involucrin) 經轉麩醯胺酶 1 (transglutaminase 1) 鈣依賴交聯形成支架;兜甲蛋白 (loricrin) 最終成為角質化細胞包膜主成分。
- 臨床關聯:轉麩醯胺酶 1 缺陷與兜甲蛋白 (loricrin) 缺陷皆可致先天性魚鱗癬 (congenital ichthyosis)。
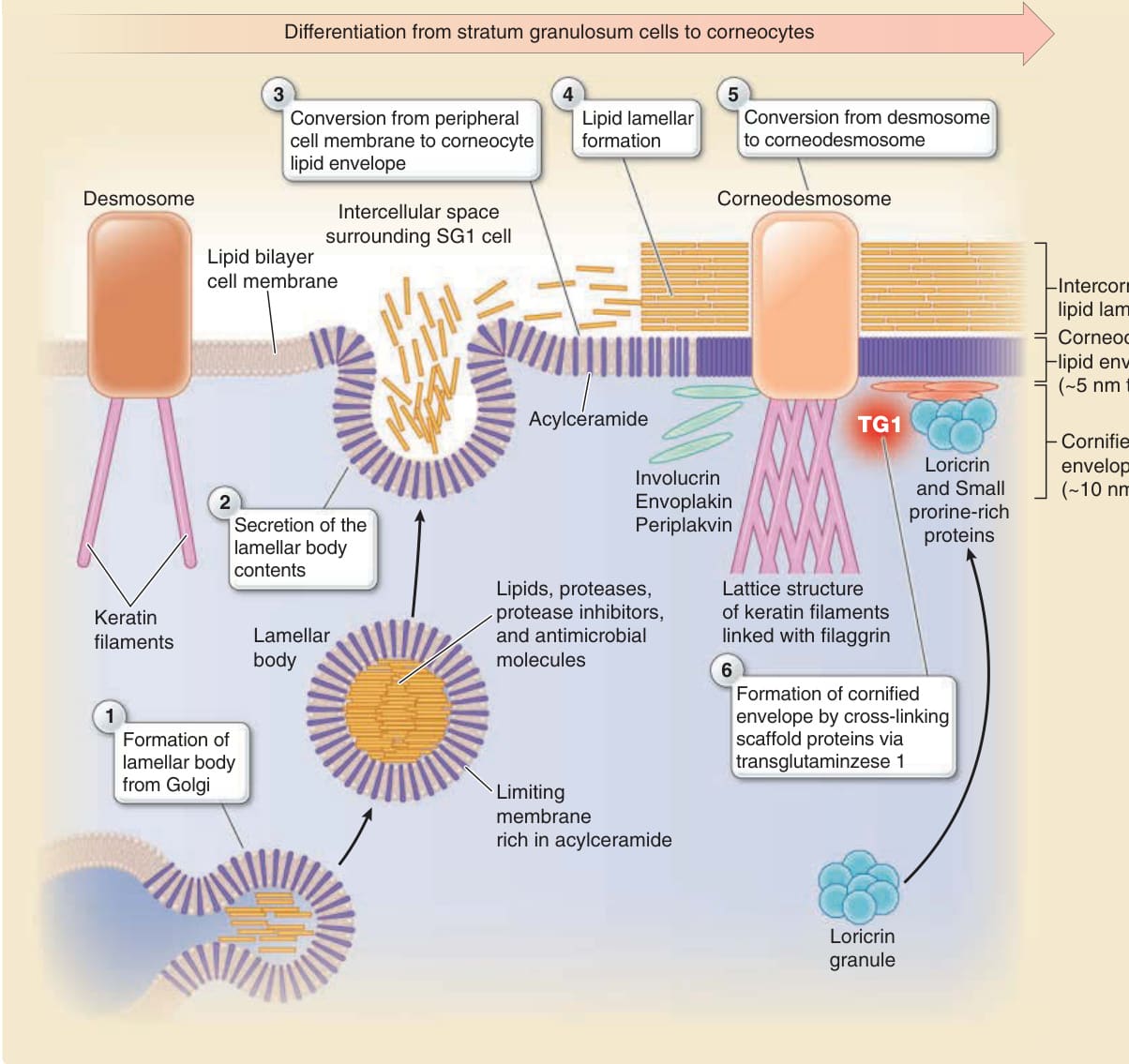
圖 14-6:角質化細胞包膜、角質細胞脂質包膜與角質細胞間脂質層板的形成
角質細胞間脂質層板與脂質組成
- 層板小體 (lamellar bodies) 由高基氏複合體產生,內含極性脂質、糖鞘脂、游離固醇與磷脂質,於 SG1/SG2 經胞吐分泌至細胞外;糖鞘脂水解生成神經醯胺 (ceramides)、磷脂質轉為游離脂肪酸 (free fatty acids),形成不透水的層板屏障。
- ABCA12 負責層板小體脂質運輸,嚴重缺陷致小丑型魚鱗癬 (Harlequin ichthyosis)。
- 角質層三大脂質類別:膽固醇 (cholesterol)、游離脂肪酸、神經醯胺。
- 膽固醇由乙醯輔酶 A 原位合成,限速步驟由 HMG-CoA 還原酶催化,屏障修復時上調;代謝中間產物 7-去氫膽固醇經 UVB 光解轉為前維生素 D,續熱異構化成維生素 D3。
- NSDHL/MSMO1 缺乏分別造成 CHILD 症候群與 MSMO1 缺乏症的發炎性魚鱗癬樣病灶(毒性中間產物累積)。
- 必需脂肪酸缺乏症 (EFAD):飲食異常或吸收不良所致,造成粗糙、脫屑、紅斑性皮膚並伴嚴重通透性屏障缺陷。
- 神經醯胺在哺乳動物全身屬次要脂質(少於膽固醇或磷脂質的 10%),但在角質層為主要脂質,以重量計佔 30% 至 40%。醯基神經醯胺由 ω-羥基神經醯胺與亞油酸 (linoleic acid) 酯化而成,如「分子鉚釘 (molecular rivet)」連結脂質雙層;其合成缺陷可致數型先天性魚鱗癬。
角質橋粒與脫屑
- 角質橋粒素 (corneodesmosin) 由層板小體分泌、整合進橋粒形成角質橋粒;角質細胞黏附取決於角質橋粒與細胞間脂質層板兩者。
- 脫屑:角質橋粒於角質層外層被降解,最外層角質細胞逐一脫落。
- 主要降解酶為激肽釋放酶 (kallikrein) 群絲胺酸蛋白酶(KLK 5、7、14),以無活性前驅物產生,由自我活化或基質裂解素 (matriptase) 活化;其活性受 LEKTI (lymphoepithelial Kazal-type-related inhibitor) pH 依賴性抑制——外層較低 pH 促進 KLK 從 LEKTI 解離、啟動降解,防止過早脫屑。
- 臨床關聯:
- 基質裂解素缺陷 → 先天性魚鱗癬(角化過度、降解受損)。
- LEKTI 缺陷 → 內瑟頓症候群 (Netherton syndrome),整層角質層易剝離(KLK 過度降解)。
- 角質橋粒素或橋粒芯蛋白 1 (desmoglein 1) 缺陷亦致整層脫落。
- 易整層脫落者傾向多種過敏(過敏原滲透增加、經皮致敏化 percutaneous sensitization)。
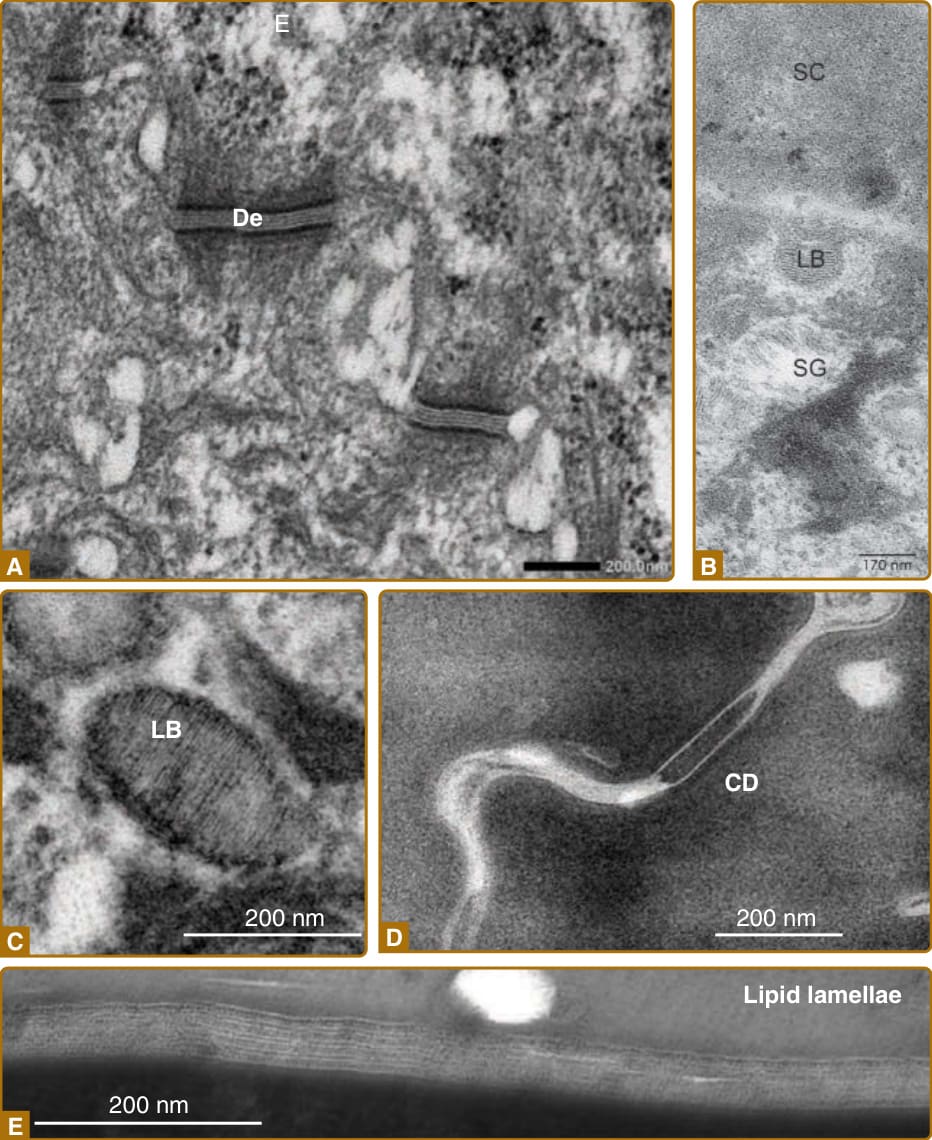
圖 14-4:角質細胞、角質橋粒與細胞間脂質層板
角蛋白細胞骨架、絲聚蛋白與天然保濕因子
- 絲聚蛋白 (filaggrin) 為角質層主要蛋白;僅表現於角質化複層上皮(皮膚有、食道/呼吸道無)。前絲聚蛋白形成透明角質顆粒 (keratohyalin granules),角質化時被處理為成熟單體,捆束角蛋白中間絲並於下層形成晶格結構 (lattice structure)。
- 角質細胞上移時,絲聚蛋白被胱天蛋白酶 14 (caspase 14)、博來黴素水解酶等降解為游離胺基酸及衍生物——如組胺酸經組胺酸酶生成反式尿刊酸 (trans-urocanic acid)、麩醯胺酸生成吡咯啶酮羧酸。
- 天然保濕因子 (natural moisturizing factors) 主要由上述胺基酸與衍生物,加乳酸、尿素、檸檬酸鹽與糖類組成。
- 臨床關聯:
- 胱天蛋白酶 14 缺乏 → 絲聚蛋白降解缺陷、魚鱗癬表型。
- 絲聚蛋白單倍劑量不足(異型合子突變)→ 尋常型魚鱗癬 (ichthyosis vulgaris)(乾燥皮膚),並傾向異位性皮膚炎 (atopic dermatitis)、食物過敏、氣喘。
- 絲聚蛋白基因突變使病人傾向異位性濕疹、過敏性鼻炎、食物過敏、併濕疹之氣喘;屏障缺陷屬中等,過敏發展另受免疫遺傳差異、皮膚微生物群與環境(低濕度、空污)調節。
角質層多區域結構與經皮吸收
- 角質層可分上、中、下三部:上層角質細胞蛋白結構被蛋白水解、如海綿般吸放水溶性小分子;中層存在通透性屏障且富含天然保濕因子、吸水保水力強;下層具角蛋白絲晶格結構,貢獻物理強度。
- 角質層為一堆「死亡」角質細胞,缺乏 ATP 依賴主動運輸、蛋白轉譯與 mRNA 轉錄等耗能活動。
- 細胞旁路徑 (paracellular pathway) 是外來分子滲透角質層的主要途徑;亦可經皮脂腺或毛囊。貼布試驗 (patch testing) 封閉性貼片影響脂質層板、增加溶質滲透。
- 小分子(如半抗原 haptens)通過角質層後可經細胞穿透途徑滲入真皮;大型肽抗原(塵蟎、卵白蛋白)難過角質層,且被緊密連接屏障阻擋。幼兒期角質層屏障缺陷與異位性皮膚炎、過敏性氣喘、食物過敏發展相關。
緊密連接
- 緊密連接由緊密連接索構成,主成分為四次跨膜蛋白閉合蛋白 (claudins)(如拉鏈鎖封閉相鄰質膜);其他成分含閉合素 (occludin)、JAM-A、三胞連蛋白、角蛋白 (angulins) 與胞內支架蛋白 ZO-1/2/3。兼具離子選擇性與大小選擇性。
- 表皮中僅 SG2 單層細胞具緊密連接並呈頂端–基底側極性;緊密連接屏障將表皮細胞外空間分為兩部分。
- 臨床關聯:閉合蛋白-1 (claudin-1) 缺乏在小鼠致緊密連接滲漏、角質層屏障形成缺陷與新生兒早期死亡;表現降低致魚鱗癬表型與皮膚發炎;人類缺乏致罕見的新生兒魚鱗癬硬化性膽管炎症候群 (neonatal ichthyosis sclerosing cholangitis syndrome)。
緊密連接與表皮內免疫細胞
- 蘭格罕細胞 (Langerhans cells):穩定狀態位於緊密連接之下、樹突向上;活化後樹突延伸超越緊密連接屏障捕捉穿透角質層的外來抗原,過程中與 SG2 角質細胞形成新緊密連接,攝取抗原而不破壞屏障;隨後遷出至引流淋巴結向 T 細胞呈現抗原。
- γ-δ T 細胞:人類表皮罕見、小鼠表皮佔比高;樹突與帶緊密連接細胞基底側對接形成免疫突觸。
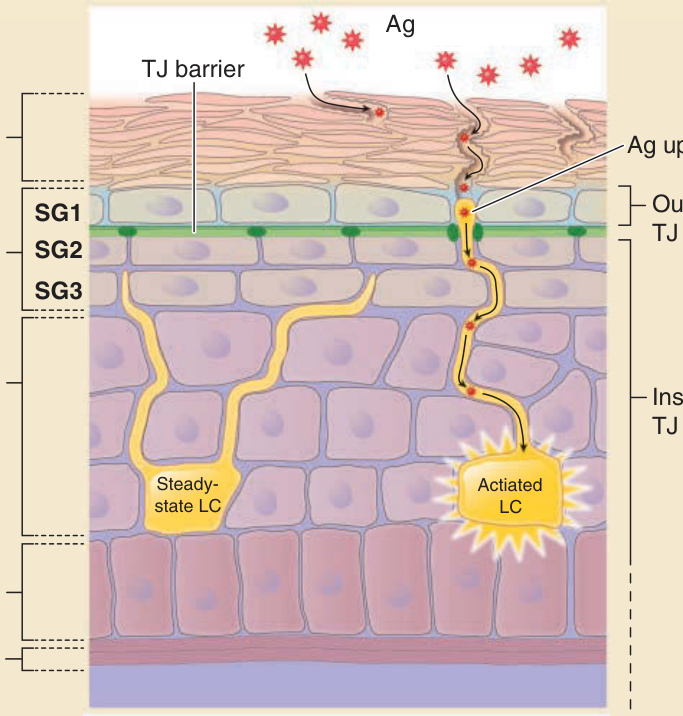
圖 14-10:角質層、緊密連接與蘭格罕細胞的空間位置
角質細胞形狀與屏障恆定
- 角質細胞依規則堆疊、相鄰細胞柱間呈鋸齒狀指狀交錯 (zigzag interdigitation);基本形狀為克耳文十四面體 (Kelvin’s tetrakaidecahedron) 的扁平變體。
- 細胞更新時,新細胞先於舊細胞下方形成新的緊密連接多邊形,暫時形成雙邊 (double-edged) 多邊形以維持屏障恆定。
抗菌屏障
- 控制皮膚微生物群以防細菌與黴菌感染。被動防護:持續脫屑更新、低碳水與低水分、弱酸性 pH(5.6 至 6.4,由游離脂肪酸、乳酸、尿刊酸及微生物代謝物造成)。
- 主動防護:抗菌蛋白 (antimicrobial proteins)/警報素,廣譜抗革蘭氏陽性與陰性菌、部分抗黴菌或抗病毒;經破壞微生物膜作用,並招募白血球修飾發炎。皮膚兩大主要抗菌蛋白為抗菌肽素 (cathelicidins) 與 β-防禦素 (β-defensins)。
- 來源:角質細胞(毛囊中更高量)、外分泌/頂泌/皮脂腺、真皮肥大細胞(儲存抗菌肽素,損傷後分泌),以及共生細菌(如表皮葡萄球菌 Staphylococcus epidermidis)。
- 人類抗菌肽素成員 LL-37 招募嗜中性球、T 細胞、單核球與肥大細胞,並經粒線體抗病毒訊號途徑促進干擾素-β 生成。
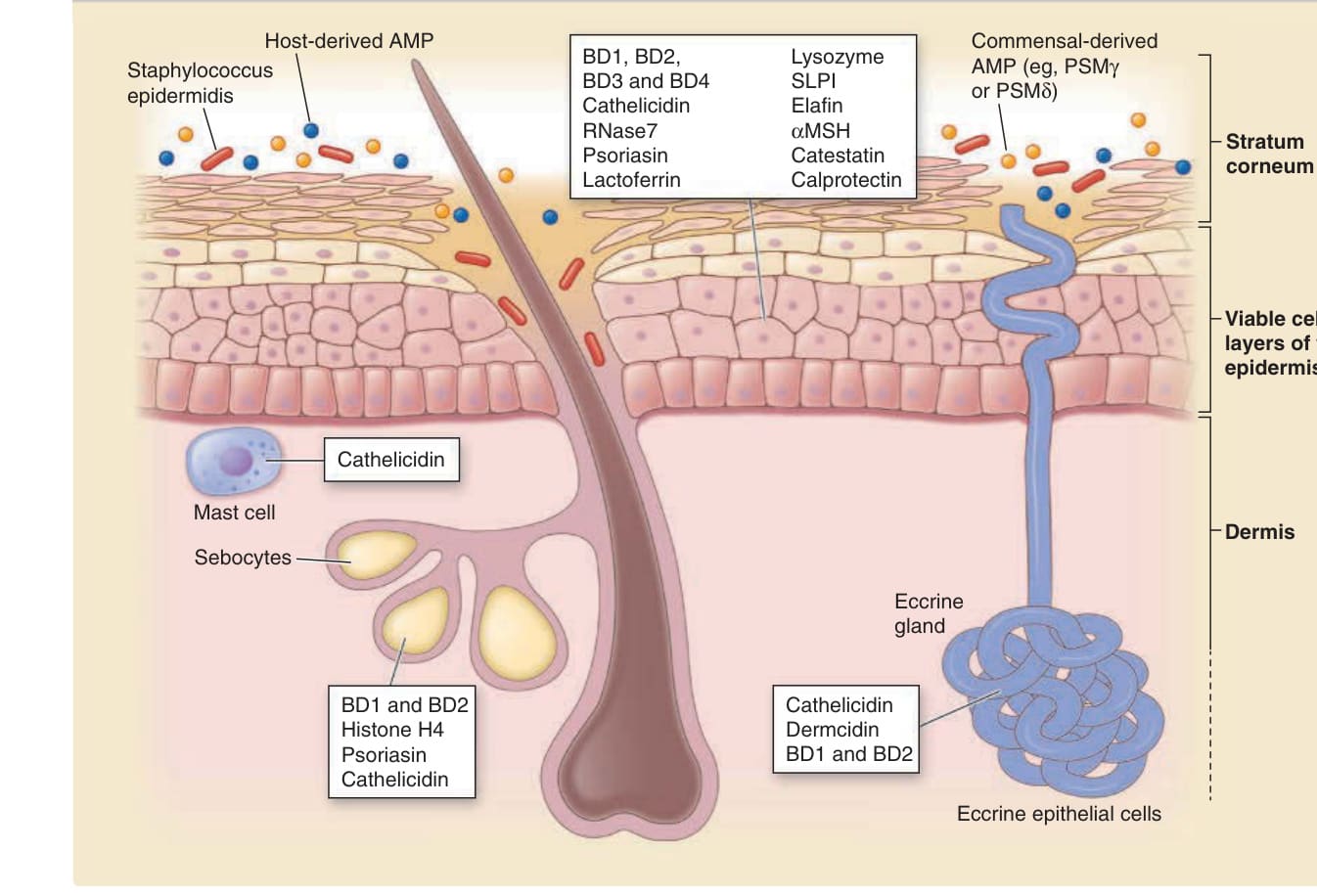
圖 14-12:產生抗菌蛋白的細胞
新生兒皮膚屏障
- 足月新生兒出生即具發育完善屏障;其角質層較薄、天然保濕因子濃度較低、經皮水分散失 (TEWL) 較大,但健康足月兒 TEWL 等於或低於成人。
- 胎脂 (vernix caseosa):第三孕期覆蓋全皮,為水、蛋白與脂質混合物,保護表皮免於羊水侵害並促進角質層形成;含溶菌酶、乳鐵蛋白,具抗菌活性。出生後保留胎脂可提高皮膚水合、降低表面 pH,提示不必如 WHO 建議立即去除。
- 早產兒屏障脆弱、真皮機械性脆弱;妊娠 28 週前缺胎脂覆蓋。妊娠 25 週分娩者僅數層角質層、TEWL 約 70 g/m²/hr,至約妊娠 35 週降至約 7 g/m²/hr;極早產兒即使產後 1 個月 TEWL 仍顯著高於足月兒。
對抗物理、紫外線與溫度應力
- 物理應力:皮膚由堅硬與柔軟組織條帶構成(堅硬角質層、柔軟角質細胞層、堅硬真皮膠原、柔軟富脂下皮)。各層強度或邊界缺陷致表皮鬆解性魚鱗癬、各型表皮鬆解水疱症、埃勒斯-當洛斯症候群。
- 紫外線:空氣–皮膚界面反射、反式尿刊酸吸收、角蛋白絲繞射限制 UV 穿透;黑色素 (melanin) 為吸收 UV、保護基因組 DNA 的主因;基底層細胞 DNA 損傷由 DNA 修復酶即時修復。
- 熱應力:主要靠外分泌汗腺 (eccrine sweat glands) 出汗蒸發冷卻;人類外分泌汗腺密度為哺乳動物中最高。無汗/少汗病人易中暑 (thermoplegia)。
- 血流控制:體溫升高時血管擴張、皮膚發紅散熱;寒冷時血管收縮、皮膚轉蒼白。動脈與靜脈緊密並列進行逆流熱交換 (countercurrent heat exchange) 以維持核心體溫。
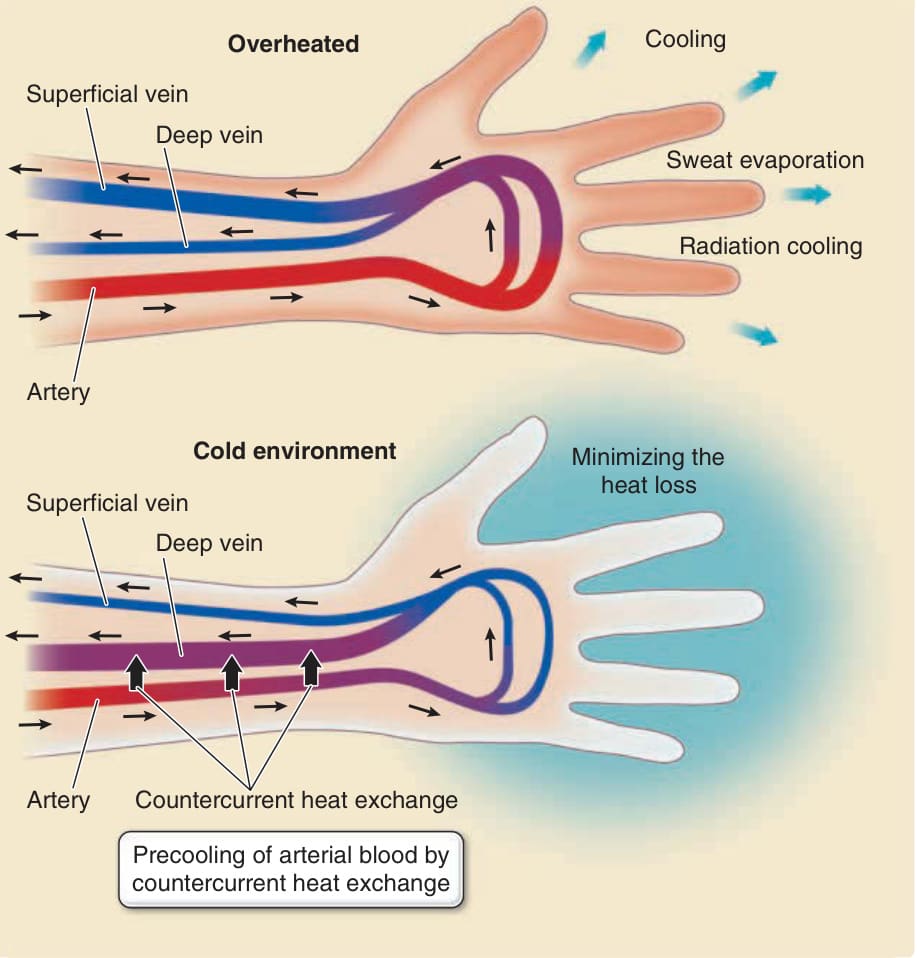
圖 14-13:對抗熱應力的脈管系統與血流控制