皮膚屏障 (Skin Barrier)
PART 2
皮膚的結構與功能 (Structure and Function of Skin)
重點一覽 (AT-A-GLANCE)
■ 皮膚最重要的功能之一,是在生物體與外在環境之間形成屏障。
■ 皮膚保護我們的身體免於乾燥 (desiccation)、物理性應力 (physical stress)、感染、過熱或散熱,以及紫外線 (ultraviolet, UV) 照射所造成的物理性損傷。
■ 皮膚由表皮 (epidermis) 所覆蓋,表皮是一層角質化的複層上皮細胞片 (cornified, stratified epithelial cellular sheet),配備有由角質層 (stratum corneum) 與緊密連接 (tight junctions) 所構成的屏障。
■ 角質層是位於體表的空氣–液體界面屏障 (air–liquid interface barrier),可防止過度的水分流失(由內向外屏障 inside–outside barrier),並防止有害物質從環境進入(由外向內屏障 outside–inside barrier)。
■ 角質層屏障由角質細胞 (corneocytes) 與角質細胞間不透水的脂質層板 (intercorneocyte water-impermeable lipid lamellae) 所組成。角質細胞被角質化細胞包膜 (cornified cell envelope) 與角質細胞脂質包膜 (corneocyte lipid envelope) 所包覆,並含有與絲聚蛋白 (filaggrin) 結合的角蛋白絲 (keratin filaments),而絲聚蛋白會被降解為天然保濕因子 (natural moisturizing factors)。
■ 緊密連接封閉了角質層下方顆粒層 (stratum granulosum) 第二層中相鄰細胞間的細胞間隙,並形成一個液體–液體界面屏障 (liquid–liquid interface barrier),限制分子經由細胞旁路徑 (paracellular pathway) 的移動。
■ 蘭格罕細胞 (Langerhans cells) 在穩定狀態下位於表皮中緊密連接屏障之下,但在活化時會將其樹突延伸至緊密連接屏障之外,於樹突末端捕捉外來抗原。
■ 抗菌肽 (antimicrobial peptides)、脂質、角質層的酸性 pH 值,以及持續的每日脫屑 (desquamation,死亡皮膚細胞的每日脫落) 控制著皮膚微生物群 (skin microbiota),並保護我們免於細菌、酵母菌、黴菌與病毒的感染。
■ 紫外線會被角質層反射,並被尿刊酸 (urocanic acid) 與黑色素 (melanin) 分子吸收,這些都能保護基因組 DNA 免於紫外線照射的損傷。
■ 出汗、血流控制,以及皮下脂肪組織 (subcutaneous adipose tissue) 中的熱儲存,保護我們免於寒冷與過熱。
前言 (INTRODUCTION)
皮膚是脊椎動物的外被 (integument)(表 14-1)。皮膚的關鍵功能之一,是在身體與外在環境之間形成一個保護性的物理屏障。分子穿越皮膚的移動受限,在哺乳動物中主要取決於表皮,尤其是角質層。表皮可防止水分、電解質、脂質與蛋白質的內外通行,亦可防止來自化學物質、細菌、黴菌、病毒、毒素與過敏原的侵害。表皮屏障形成的缺陷會造成各種先天性疾病(表 14-2)。皮膚保護身體免於各種外在應力。在保護身體免於熱與冷的應力方面,皮膚透過調節血流、汗液生成、脂肪層的熱儲存,以及棕色脂肪細胞 (brown fat cells) 的產熱作用,將內臟器官維持在一定的恆定溫度。皮膚透過協調角質層堅硬的表面盔甲、角蛋白細胞骨架 (keratin cytoskeleton),以及角質細胞之間的細胞黏附、表皮–真皮交界處 (epidermal–dermal junctions) 與真皮膠原纖維 (dermal collagen fibers),來保護身體免於物理性應力;這些將於第 15 章與第 60 章中討論。此外,表皮反射並吸收來自太陽的紫外線 (ultraviolet, UV) 輻射,以保護細胞的基因組 DNA;這在預防癌化 (carcinogenesis) 方面特別重要,將於第 19 章與第 20 章中討論。皮膚附屬器 (skin appendages),如汗腺與皮脂腺、毛髮與指甲,也具有物理與化學屏障功能;這些分別於第 6、7、8 章中討論。皮膚也配備有對先天性與後天性免疫系統的免疫屏障。抗菌蛋白 (antimicrobial proteins) 是一群多樣化的蛋白質,在表皮表面形成對抗微生物的化學屏障。角質細胞上的類鐸受體 (Toll-like receptors) 偵測病原體相關分子模式 (pathogen-associated molecular patterns),例如細菌廣泛共有但可與宿主分子區分的脂蛋白與肽聚糖 (peptidoglycans),並控制對抗微生物的免疫反應。皮膚內的樹突細胞 (dendritic cells) 主宰對外來侵害的後天性免疫與過敏反應。保護我們皮膚的後天性免疫機轉將於第 10 章與第 11 章中討論。
外被 (Integument) 動物或植物堅韌的外層保護層,將身體與外在環境劃分開來。
封閉性連接 (Occlusive junction) 封閉細胞間隙並控制細胞旁分子移動的細胞–細胞黏附性連接。無脊椎動物與脊椎動物具有不同的封閉性連接結構。無脊椎動物的封閉性連接稱為隔膜連接 (septate junction),脊椎動物的則稱為緊密連接 (tight junction)。
比較生物學 (Comparative Biology) 比較生物學利用物種或分類群之間的自然變異與差異,藉由研究系統發育 (phylogenies)、演化、發育、生理學與基因組學,來理解生命的模式。
克耳文十四面體 (Kelvin’s Tetrakaidecahedron) 由八個六邊形與六個正方形組成的十四面體,由 William Thomson(克耳文勳爵 Lord Kelvin)於 1887 年提出,作為以每個細胞最小表面積將等大細胞堆疊在一起以填滿空間的最佳形狀。
經皮水分散失 (Transepidermal Water Loss) 水分經由表皮從體內傳向體外的流失。經皮水分散失的測量有助於評估角質層屏障,因為經皮水分散失的升高率基本上與屏障損傷或形成的程度成正比。
表皮屏障的比較生物學 (COMPARATIVE BIOLOGY OF THE EPIDERMAL BARRIER)
脂質雙層細胞膜 (lipid bilayer cell membrane) 在單細胞生物中提供了一個基本屏障。細胞膜將細胞與外在環境區隔開來。細胞的恆定 (homeostasis) 以耗能 (energy-dependent) 的方式維持在由細胞膜所劃分的隔室內。由於脂質雙層膜極為脆弱,多數單細胞生物在細胞膜之外具有一個額外的屏障結構(即細胞壁 cell wall),其功能如同一種盔甲(圖 14-1)。多細胞生物身體的屏障類似於其對單細胞生物的屏障。單細胞生物的細胞膜對應於表皮,表皮是劃定多細胞生物表面的上皮細胞片(見圖 14-1)。要以一層細胞片形成屏障,防止水分與溶質經由細胞間隙滲漏至關重要。因此,細胞間隙由封閉性連接所封閉,例如節肢動物的隔膜連接與脊椎動物的緊密連接(見表 14-1)。表皮在頭索動物 (cephalochordates) 與尾索動物 (urochordates) 中由單層細胞組成,但在脊椎動物中則為多層(複層 stratified)細胞(見圖 14-1)。由於表皮細胞在直接暴露於嚴苛的外在環境因子(例如低張的淡水、高張的海水或乾燥空氣)時相當脆弱,較高等生物的表皮表面被各種外部屏障所覆蓋。這些屏障的例子包括節肢動物的角質層 (cuticles)、被囊類動物 (tunicates) 的被囊 (tunics)、魚類與兩棲類的黏液 (mucous),以及成年兩棲類、爬蟲類、鳥類與哺乳類的角質化細胞層(角質層 stratum corneum)(見圖 14-1)。¹
表皮的基本結構 (BASIC STRUCTURE OF THE EPIDERMIS)
人類表皮是一層複層上皮細胞片,其最上部角質化形成角質層。表皮中具活性(去核 enucleated)的細胞層由基底層(stratum basale,基底細胞層)、棘層(stratum spinosum,棘細胞層)與顆粒層(stratum granulosum,顆粒細胞層)所組成(圖 14-2)。顆粒層由至少三層扁平的顆粒細胞組成。²⁻⁵ 由外而內,這些層分別命名為 SG1、SG2 與 SG3,而緊密連接封閉 SG2 層的細胞間隙。³,⁵ 角質層由死亡的角質化細胞(角質細胞 corneocytes)所組成,並作為一個空氣–液體界面屏障。角質層的細胞間隙被不透水的脂質層板所填滿。在角質層之下,細胞浸潤於水環境中。表皮的細胞外水環境被緊密連接屏障(一個液體–液體界面屏障)分為兩部分(見圖 14-2)。為形成角質層,SG1 細胞藉由以脂質層板填滿其細胞間隙而終末分化 (terminally differentiate) 為角質細胞。表皮的多層結構得以維持,而表皮細胞持續更新。表皮細胞僅在基底層增生,隨著與基底膜 (basement membrane) 分離而分化,向上移動,在顆粒層變得扁平,於 SG2 層形成緊密連接,在 SG1 層失去緊密連接,終末分化為角質細胞,並以鱗狀片屑 (squamous scales) 的形式從角質層頂部脫落。表皮的持續更新使得附著於皮膚的外來物質能隨著每日的片屑脫落而被丟棄。
角質層 (STRATUM CORNEUM)
人類皮膚的角質層是身體表面最外層的屏障。人類的角質層厚度約為 10 至 20 µm,並含有約 10 至 25 層角質化細胞。⁶ 角質層直接暴露於空氣中,並保護內層細胞免於乾燥損傷。它作為一個雙向屏障,不僅防止外來分子與微生物向內滲透,也防止水分與溶質向外滲漏。
角質層的基本結構 (BASIC STRUCTURE OF THE STRATUM CORNEUM)
角質層由複層的角質細胞與角質細胞間富含脂質的基質 (lipid-rich matrix) 所組成(圖 14-3)。角質層的屏障功能取決於角質細胞中富含蛋白質的物質,以及細胞間富含脂質的基質兩者。角質細胞是終末分化的死亡角質細胞 (keratinocytes),藉由稱為角質橋粒 (corneodesmosomes) 的蛋白質性細胞–細胞黏附複合體,以及細胞間脂質層板的黏附力,彼此相互黏附(圖 14-4)。在以蘇木精與伊紅 (hematoxylin and eosin) 對石蠟包埋皮膚切片進行的常規染色中,角質層呈現一種籃網狀結構 (basket-weave structure),這可能使角質層被誤認為是一個不具屏障功能的多孔結構;事實上,籃網狀結構是檢體處理過程中的人為假象 (artifact)。在使用二甲苯 (xylene) 去除石蠟的過程中,角質細胞間的脂質被萃取出來,角質細胞之間依賴細胞間脂質的黏附力遂被削弱。由於角質橋粒使角質細胞之間的側向黏附比頂底向 (apicobasal) 黏附更為穩定,角質細胞層在維持其側向黏附的同時彼此分離,導致石蠟包埋切片中籃網狀結構的形成⁷,⁸(圖 14-5)。適當的固定與染色程序則能揭露皮膚垂直切片中角質層緊密堆積、組織良好的結構²,⁹(見圖 14-5)。
角質化細胞包膜與角質細胞脂質包膜的形成 (FORMATION OF A CORNIFIED CELL ENVELOPE AND CORNEOCYTE LIPID ENVELOPE)
顆粒層細胞終末分化為角質細胞以形成角質層。在角質化過程中,一層由高度交聯 (highly cross-linked) 的不溶性蛋白質所構成、厚約 10 nm 的角質化細胞包膜,在質膜 (plasma membrane) 之下形成¹⁰,而質膜的脂質雙層被一層厚約 5 nm 的醯基神經醯胺 (acylceramides) 所取代,此即稱為角質細胞脂質包膜¹¹⁻¹³(圖 14-6)。角質化細胞包膜與角質細胞脂質包膜的形成描述如下¹⁴,¹⁵(見圖 14-6)。表現於顆粒層細胞中的兜膜蛋白 (envoplakin)、外周斑蛋白 (periplakin) 與兜甲蛋白 (involucrin) 以鈣依賴 (calcium-dependent) 的方式與質膜內表面結合,並由轉麩醯胺酶 1 (transglutaminase 1) 彼此交聯。¹⁶ 轉麩醯胺酶 1 也與其他膜相關蛋白與橋粒蛋白 (desmosomal proteins) 交聯,將細胞連接與相關的細胞骨架固定於蛋白質性的支架上。以兜膜蛋白–外周斑蛋白–兜甲蛋白為基礎的支架最終沿著質膜的整個內表面形成一個單分子層。¹⁰,¹⁷⁻¹⁹
已知轉麩醯胺酶 1 的缺陷會造成先天性魚鱗癬(congenital ichthyosis)(見表 14-2)。²⁰
在支架覆蓋質膜內表面之前,層板小體 (lamellar bodies) 由高基氏複合體 (Golgi complex) 產生並與質膜融合。層板小體的界膜 (limiting membrane) 富含由 ω-羥基化超長鏈脂肪酸 (omega-hydroxylated ultra-long-chain fatty acids) 所組成的醯基神經醯胺。界膜與質膜的融合逐漸增加質膜脂質雙層內醯基神經醯胺的量。醯基神經醯胺藉由膜錨定型的轉麩醯胺酶 1,與角質化包膜支架的外表面共價結合。²¹,²² 醯基神經醯胺最終取代質膜,此即稱為角質細胞脂質包膜¹¹,¹²(見圖 14-6)。以兜膜蛋白–外周斑蛋白–兜甲蛋白為基礎的支架在角質化的後期階段進一步被強化。在支架的內表面,兜甲蛋白分子從細胞質中轉位,並共價交聯至支架上以構建角質化細胞包膜。兜甲蛋白是一種可能有助於角質化細胞包膜抗水性的不溶性蛋白,最終成為角質化細胞包膜的主要成分。不同量的小分子富含脯胺酸蛋白 (small proline-rich proteins),連同少量的其他蛋白質(如重複蛋白 repetin、毛透明蛋白 trichohyalin、胱抑素 A cystatin A、彈性蛋白酶抑制因子 elafin 與晚期包膜蛋白 late envelope proteins),也交聯至角質化包膜的內表面。²³ 已知兜甲蛋白的缺陷會造成先天性魚鱗癬(見表 14-2)。²⁴
角質細胞間脂質層板的形成 (FORMATION OF THE LAMELLAE OF INTERCORNEOCYTE LIPIDS)
顆粒層的角質細胞發展出一種特定的層板小體系統,使角質細胞間脂質層板得以分泌。²⁵ 層板小體由高基氏複合體產生,並以細胞內囊泡 (intracellular vesicles) 的形式儲存於 SG3 細胞的細胞質中。層板小體富含極性脂質 (polar lipids)、糖鞘脂 (glycosphingolipids)、游離固醇 (free sterols) 與磷脂質 (phospholipids)。ABCA12 在層板小體界膜上的細胞脂質運輸 (lipid trafficking) 中發揮功能,其嚴重缺陷會造成小丑型魚鱗癬(Harlequin ichthyosis)(見表 14-2)。²⁶,²⁷ 層板小體也可能含有蛋白質,例如修飾脂質的水解酶、修飾角質橋粒的角質橋粒素 (corneodesmosins)、抗菌肽,以及控制脫屑的蛋白酶與蛋白酶抑制劑。層板小體的內容物經由頂端細胞膜 (apical cell membrane) 的胞吐作用 (exocytosis) 被輸送至細胞外環境,最可能發生於 SG2 細胞與 SG1 細胞,以填滿圍繞 SG1 細胞的細胞外空間²⁸(見圖 14-2)。
從層板小體胞吐出的脂質,隨後在 SG1 細胞角質化的過程中被組織成一種特徵性的、與角質化細胞包膜平行排列的層板結構(見圖 14-4)。角質細胞脂質包膜中共價結合的醯基神經醯胺作為一個支架,使細胞間脂質得以規則地形成層板(見圖 14-4 與 14-6)。²² 在層板小體被擠出至細胞外空間後,極性脂質被酵素轉化為非極性產物。糖鞘脂被水解以生成神經醯胺 (ceramides),而磷脂質則被轉化為游離脂肪酸 (free fatty acids)。這些脂質形成複層脂質雙層的細胞間層板成分,是一種緊密堆積於角質細胞間隙中的極為緻密的結構,從而形成不透水的屏障。
角質層的脂質組成 (LIPID COMPOSITION OF THE STRATUM CORNEUM)
細胞間脂質對於角質層通透性屏障 (permeability barrier) 的形成不可或缺。角質層中的主要脂質類別為膽固醇 (cholesterol)、游離脂肪酸與神經醯胺。²⁹
膽固醇構成表皮中具活性細胞層質膜的一部分,以及角質層中細胞間脂質層板的一部分。雖然基底層的角質細胞能夠從循環中再吸收膽固醇,表皮角質細胞會主動生物合成膽固醇與游離脂肪酸(圖 14-7)。表皮中大部分的膽固醇是由乙醯輔酶 A(acetyl coenzyme A, acetyl CoA)原位 (in situ) 合成。³⁰ 膽固醇生物合成的限速步驟由羥甲基戊二醯輔酶 A 還原酶(hydroxymethylglutaryl CoA [HMG CoA] reductase)所催化(見圖 14-7)。表皮膽固醇的生物合成在屏障修復 (barrier repair) 期間上調。³¹ 表皮中的膽固醇生物合成途徑對於維生素 D 的生成也很重要。³²
膽固醇生物合成的代謝中間產物 7-去氫膽固醇 (7-dehydrocholesterol),在 UVB 輻射的光解反應 (photolytic reaction) 中被轉化為前維生素 D (previtamin D);隨後經由熱異構化 (thermal isomerization) 形成維生素 D3。對應於膽固醇生物合成的 NSDHL 與 MSMO1 酵素缺乏,會分別透過毒性中間產物的累積,造成 CHILD 症候群(congenital hemidysplasia with ichthyosiform erythroderma and limb defects,伴有魚鱗癬樣紅皮症與肢體缺陷的先天性偏側發育不全)與 MSMO1 缺乏症中的發炎性魚鱗癬樣皮膚病灶³³⁻³⁵(見表 14-2)。皮膚含有游離脂肪酸,以及結合於三酸甘油酯 (triglycerides)、磷脂質、糖基神經醯胺 (glycosylceramides) 與神經醯胺中的脂肪酸。雖然膽固醇的生物合成源自 HMG-CoA,游離脂肪酸的合成則源自由乙醯輔酶 A 經乙醯輔酶 A 羧化酶 (acetyl-CoA carboxylase) 所產生的丙二醯輔酶 A (malonyl-CoA)(見圖 14-7)。飽和與單元不飽和脂肪酸 (monounsaturated fatty acids) 在表皮中合成。然而,並非所有脂肪酸都能由表皮或人體合成。人體無法生成的脂肪酸稱為必需脂肪酸 (essential fatty acids)。必需脂肪酸缺乏症(essential fatty acid deficiency, EFAD),由人類異常的飲食或吸收不良所造成,或在囓齒類中以實驗誘導,會導致粗糙、脫屑與紅斑性的皮膚,並伴有嚴重的表皮通透性屏障缺陷。³⁶,³⁷
神經醯胺由稱為鞘氨醇鹼基 (sphingoid bases) 的長鏈胺基醇 (long-chain amino alcohols) 經由醯胺鍵 (amide bond) 與脂肪酸連接所組成(見圖 14-7)。神經醯胺由絲胺酸棕櫚醯轉移酶 (serine palmitoyltransferase) 合成,並由表皮中葡萄糖基神經醯胺 (glucosylceramide) 與鞘磷脂 (sphingomyelin) 兩者的水解所合成。雖然神經醯胺在哺乳動物體內是一種次要的脂質成分(少於膽固醇或磷脂質的 10%),它在角質層中卻是主要的脂質成分,以重量計佔脂質的 30% 至 40%。²⁹ 如此高的神經醯胺含量僅見於角質層,而不見於顆粒層、棘層或基底層。此種分布顯示神經醯胺的生物合成受到時空調控 (spatiotemporally controlled),並在終末分化為角質細胞的最上層角質細胞中高度活化。在分子異質性顯著的表皮神經醯胺中,醯基神經醯胺不僅對如前所述的角質細胞脂質包膜的形成不可或缺,對角質細胞間脂質層板的適當組織以及角質層的屏障功能也是如此。³⁸⁻⁴² 醯基神經醯胺經由 ω-羥基神經醯胺 (omega-hydroxyceramide) 與亞油酸 (linoleic acid) 的酯化作用所合成。醯基神經醯胺是一種不尋常的神經醯胺,其 N-醯基鏈 (N-acyl chain) 由 ω-羥基化超長鏈脂肪酸所組成(見圖 14-7)。已有人提出,醯基神經醯胺的超長鏈脂肪酸跨越脂質層板的一層雙層,將脂質層板中的兩層膜連結在一起,從而作為一種分子鉚釘 (molecular rivet) 以形成複層脂質層板。⁴³ 醯基神經醯胺生物合成途徑的缺陷會造成數種類型的先天性魚鱗癬⁴⁴(見表 14-2)。
角質橋粒 (CORNEODESMOSOMES)
橋粒 (Desmosomes) 是顆粒層、棘層與基底細胞層中主要的細胞–細胞黏附結構。角質橋粒素由層板小體分泌至圍繞 SG1 細胞的細胞外空間(見圖 14-2 與 14-6),然後整合進橋粒,導致角質橋粒(即角質細胞之間特定的細胞黏附結構)的形成⁴⁵(見圖 14-4)。角質細胞之間的黏附取決於角質橋粒與細胞間脂質層板兩者。角質橋粒在角質層的外層被降解,最外層的角質細胞被一個接一個地從角質層的頂層脫落;此過程稱為脫屑 (desquamation)。⁴⁶
涉及角質橋粒降解的主要蛋白酶,是屬於激肽釋放酶 (kallikrein) 群的絲胺酸蛋白酶 (serine proteases)。已知激肽釋放酶 5、7 與 14 存在於角質層中。這些激肽釋放酶也由層板小體分泌至圍繞 SG1 細胞的細胞外空間(見圖 14-2)。激肽釋放酶以無活性的前驅物形式產生,並經由激肽釋放酶本身(自我活化 autoactivation)或基質裂解素 (matriptase,一種跨膜絲胺酸蛋白酶) 的蛋白水解轉化而活化。⁴⁷,⁴⁸ 活化的激肽釋放酶的蛋白酶活性被認為受到淋巴上皮 Kazal 型相關抑制劑(lymphoepithelial Kazal-type-related inhibitor, LEKTI)直接結合的抑制,LEKTI 也經由層板小體分泌至細胞間隙中。⁴⁹ 激肽釋放酶以無活性前驅物形式產生,以及其蛋白酶活性被 LEKTI 所抑制,被認為能禁止角質層下層中角質橋粒的降解,從而防止過早的脫屑。⁵⁰ LEKTI 與激肽釋放酶的結合已被證明是 pH 依賴的。角質層外層細胞間隙較低的 pH 值被認為能促進激肽釋放酶從 LEKTI 解離,以及激肽釋放酶依賴的角質橋粒降解(見圖 14-2)。⁵¹⁻⁵³ 最外層的角質細胞最終從皮膚脫落。
基質裂解素的先天性缺陷會誘發先天性魚鱗癬,表現為角化過度 (hyperkeratosis) 與角質橋粒降解受損,可能是由激肽釋放酶蛋白酶活化不足所造成。⁵⁴ 相對地,LEKTI 的先天性缺陷會誘發內瑟頓症候群(Netherton syndrome),其中整層角質層容易被剝離,可能是由激肽釋放酶對角質橋粒的降解增強所造成。⁵⁵ 此種整層角質層的脫落也見於角質橋粒素或橋粒芯蛋白 1(desmoglein 1)的先天性缺陷⁵⁶⁻⁵⁸(見表 14-2)。容易發生角質層整層脫落的病人,傾向發展出各種過敏性病況,可能是經由過敏原透過有缺陷的皮膚屏障增加滲透,以及促進經皮致敏化 (percutaneous sensitization)。⁵,⁵⁹
角蛋白細胞骨架與絲聚蛋白 (KERATIN CYTOSKELETON AND FILAGGRIN)
絲聚蛋白是角質層的主要蛋白質。絲聚蛋白基因中的各種突變使病人傾向發展出異位性濕疹 (atopic eczema)、過敏性鼻炎、食物過敏,以及併發濕疹的氣喘。⁶⁰⁻⁶³
在人類中,絲聚蛋白僅表現於角質化的複層上皮;因此,它表現於皮膚,但不表現於食道或呼吸道。⁶⁴ 前絲聚蛋白 (Profilaggrin) 表現於顆粒層,並形成稱為透明角質顆粒 (keratohyalin granules) 的細胞內聚集體。在角質化過程中,前絲聚蛋白被各種蛋白酶處理為成熟的絲聚蛋白單體⁶³(圖 14-8)。絲聚蛋白單體捆束角蛋白中間絲 (keratin intermediate filaments),並可能在角質層的下層形成一種晶格結構 (lattice structure)。⁶⁵,⁶⁶
此晶格結構可在電子顯微鏡下觀察到。在角質化的後期階段,與絲聚蛋白捆束在一起的角蛋白絲與角質化細胞包膜交聯。死亡的角質化細胞失去其大部分的細胞內胞器,主要由共價附著於並包被於細胞包膜內的捆束中間絲所組成。
絲聚蛋白降解與天然保濕因子 (FILAGGRIN DEGRADATION AND NATURAL MOISTURIZING FACTORS)
絲聚蛋白單體在角質層最內層捆束角蛋白絲。由於新的角質細胞持續在底部供應,而舊的角質細胞從角質層表面脫屑,每個角質細胞逐漸向上層移動。在角質細胞向上移動的過程中,細胞內的絲聚蛋白分子被蛋白酶(包括胱天蛋白酶 14 [caspase 14] 與博來黴素水解酶 [bleomycin hydrolase])降解為游離胺基酸及其衍生物。這些衍生物的代表性例子,包括由組胺酸 (histidine) 經組胺酸酶 (histidase) 催化而成的反式尿刊酸 (trans-urocanic acid),以及由麩醯胺酸 (glutamine) 催化而成的吡咯啶酮羧酸 (pyrrolidone carboxylic acid)。⁶³,⁶⁷⁻⁶⁹
角質層的天然保濕因子主要由這些胺基酸及其衍生物,連同乳酸 (lactic acid)、尿素 (urea)、檸檬酸鹽 (citrate) 與糖類所組成。⁶¹,⁷⁰ 胱天蛋白酶 14 的缺乏會造成絲聚蛋白適當降解的缺陷,並導致魚鱗癬的表型⁷¹(見表 14-2)。絲聚蛋白的單倍劑量不足 (Haploinsufficiency),因編碼絲聚蛋白的基因中各種異型合子突變 (heterozygous mutations) 所致,會造成尋常型魚鱗癬 (ichthyosis vulgaris)(一種乾燥皮膚的病況)⁷²,並使病人傾向發展出異位性皮膚炎 (atopic dermatitis) 以及伴隨濕疹的各種過敏性疾病(如食物過敏與氣喘)。⁷³,⁷⁴(見表 14-2)。絲聚蛋白的減少會誘發天然保濕因子的減少、角質層最內層角蛋白絲晶格結構的擾亂,以及角質層屏障破損發生率可能的增加。⁷⁵ 角質層屏障的此種脆弱可能增加外來過敏原滲透進入皮膚的機會,以及對各種過敏原經皮致敏化的風險。多數患有嚴重角質層屏障缺陷的病人,例如內瑟頓症候群的病人,都有多重過敏。相對地,多數具有絲聚蛋白基因突變的病人基本上發展出尋常型魚鱗癬,而僅有部分這些病人發展出過敏性疾病。這顯示絲聚蛋白突變所誘發的屏障缺陷是中等程度的,而這些病人過敏性疾病的發展是由各種額外因素所調節,例如免疫反應的遺傳差異⁷⁶、皮膚微生物群的差異⁷⁷⁻⁷⁹,以及環境因素(如低空氣濕度⁸⁰與空氣污染⁸¹)。
角質層的多區域結構 (MULTIDOMAIN STRUCTURE OF THE STRATUM CORNEUM)
由於共價交聯蛋白質的不溶性,對角質層進行傳統的細胞生物學分析一直受到阻礙。為克服此困難,角質層已透過各種特殊方法進行分析,例如冷凍電子顯微鏡 (cryoelectron microscopy)、免疫電子顯微鏡 (immunoelectron microscopy)、拉曼光譜顯微分析 (Raman spectromicroscopy analysis),以及飛行時間二次離子質譜分析 (time-of-flight secondary ion mass spectroscopy)。⁸²⁻⁸⁵ 根據這些分析的結果,角質層被認為可分為幾個具不同特性的不同部分,最可能分為三部分(上層、中層與下層)。例如,水溶性小分子在角質層的上部浸入與滲出,該處角質細胞以蛋白質為基礎的結構被蛋白水解降解,角質細胞被認為發揮如同海綿的功能。⁸³,⁸⁵ 水溶性小分子進一步滲透進入角質層中部則相當受限,顯示此結構的中部存在通透性屏障。⁸³ 角質層的中部也具有吸收與保持水分的巨大潛能,可能對應於天然保濕因子的富集,而天然保濕因子大多由成熟絲聚蛋白的降解所產生。⁶³,⁸² 角質層下部的角質細胞顯示由成熟絲聚蛋白單體所整合的角蛋白絲細胞內晶格結構,被認為有助於角質層的物理強度⁸⁴(圖 14-9)。角質層是一堆「死亡」的角質細胞,缺乏任何耗能的活動,例如腺苷三磷酸(adenosine triphosphate)依賴的主動運輸、蛋白質的轉譯,以及 mRNA 的轉錄。角質層如何利用其上層、中層與下層的不同特徵來維持恆定,目前仍大多未知。
經由角質層的經皮吸收 (PERCUTANEOUS ABSORPTION THROUGH THE STRATUM CORNEUM)
了解外來分子如何滲透表皮,不僅對於更好地理解各種皮膚疾病的致病機轉很重要,對於局部治療的藥物傳遞 (drug delivery) 的發展也很重要。細胞旁路徑是外來分子能滲透角質層的主要途徑(見圖 14-3)。其他進入皮膚的滲透途徑為經由皮脂腺或毛囊,但這些途徑的確切機轉仍有待闡明。⁸⁶
貼布試驗 (patch testing) 中所使用的封閉性貼片會影響角質細胞間脂質層板,並增加溶質與溶劑經由角質層的滲透。⁸⁷ 在通過角質層後,小分子(如半抗原 haptens)可能經由細胞穿透途徑 (transcellular pathway) 輕易地滲透真皮(見圖 14-9)。相對地,大型肽抗原(如塵蟎或卵白蛋白 [egg albumin] 的抗原),不僅難以通過角質層屏障,也被緊密連接屏障禁止進一步滲透進入皮膚。近期研究揭示,幼兒期角質層屏障的缺陷與各種過敏性疾病(如異位性皮膚炎、過敏性氣喘與食物過敏)的發展有關,可能是經由抗原透過角質層增加滲透以及經皮致敏化。⁷⁴,⁸⁸⁻⁹¹
緊密連接 (TIGHT JUNCTIONS)
多細胞生物利用上皮細胞片將其身體與外在環境分隔,或將其器官彼此分隔。上皮作為對化學組成不同的液體的選擇性通透屏障 (selective permeability barriers)(圖 14-10)。封閉性連接是特化的細胞間黏附複合體,對於限制分子經由細胞旁路徑的移動至關重要(見圖 14-10),因為它們封閉了上皮細胞片的細胞間隙。⁹² 緊密連接在脊椎動物中促成此屏障。例如,在人類的腸道中,配備有緊密連接屏障的腸上皮將體液與腸腔的液體分隔開來。
緊密連接的結構 (STRUCTURE OF TIGHT JUNCTIONS)
在單層上皮中,緊密連接位於頂端連接複合體(apical junctional complex)的最頂端。⁹³(見圖 14-9)。頂端連接複合體由緊密連接、帶狀的黏著連接 (beltlike adherens junctions) 與橋粒所組成。細胞膜在緊密連接處被分為兩部分——頂端細胞膜與基底側細胞膜 (basolateral cell membrane)——兩者具有不同的膜蛋白組成。緊密連接由緊密連接索 (tight junction strands) 所組成,緊密連接索主要由質膜上的四次跨膜蛋白閉合蛋白 (claudins) 所構成,可能發揮如同拉鏈鎖 (zip lock) 的功能以封閉相鄰的質膜,從而形成對抗分子經由細胞旁路徑移動的屏障(見圖 14-9)。⁹⁴,⁹⁵ 緊密連接的其他主要成分為跨膜蛋白閉合素 (occludin)、連接黏附分子 A(junctional adhesion molecule A, JAM-A)、三胞連蛋白 (tricellulin) 與角蛋白 (angulins),以及細胞內支架蛋白 ZO-1、2 與 3。⁹⁶,⁹⁷ 緊密連接不僅形成一個不可滲透的屏障,也作為離子選擇性與大小選擇性的屏障 (ion- and size-selective barriers),其緊密程度因細胞類型及其結構性黏附分子的組成而異。⁹⁷⁻⁹⁹
緊密連接的功能 (FUNCTION OF TIGHT JUNCTIONS)
對正常人類軀幹皮膚以及小鼠耳部、腹部與背部皮膚的觀察揭示,表皮中僅有 SG2 層的單層細胞配備有緊密連接,並顯示頂端與基底側細胞膜極性(見圖 14-2 與 14-9)。¹⁰⁰,¹⁰¹ 表皮的細胞外空間被緊密連接屏障分為兩部分。⁵ SG2 與 SG1 細胞經由其頂端細胞膜的層板小體分泌,將各種分子(即脂質與蛋白酶)分泌至圍繞 SG1 細胞的細胞外空間,這些細胞外空間位於緊密連接屏障之外,並可能與緊密連接屏障內側的環境區隔開來(見圖 14-2)。閉合蛋白-1(claudin-1,表皮主要的閉合蛋白)的缺乏會在小鼠中造成表皮緊密連接屏障的滲漏,以及角質層屏障形成的缺陷,導致新生兒早期死亡(可能由體表的水分流失所造成)。¹⁰²
小鼠中閉合蛋白-1 表現的降低會誘發魚鱗癬表型與皮膚發炎。¹⁰³ 人類中閉合蛋白-1 的缺乏會導致一種極為罕見的先天性疾病,稱為新生兒魚鱗癬硬化性膽管炎症候群(neonatal ichthyosis sclerosing cholangitis syndrome)¹⁰⁴(見表 14-2)。這些觀察顯示,一個功能性的緊密連接屏障對於角質層屏障的適當形成以及表皮恆定的維持至關重要。
緊密連接屏障與表皮內免疫介導細胞之間的交互作用 (INTERACTION BETWEEN TIGHT JUNCTION BARRIERS AND INTRAEPIDERMAL IMMUNE-MEDIATED CELLS)
蘭格罕細胞是存在於表皮中、具有多個樹突的抗原呈現細胞。在人類與小鼠表皮中,它們在穩定狀態下停留於緊密連接之下,並將其樹突向上投射。¹⁰¹,¹⁰⁵ 在活化之後,蘭格罕細胞將其樹突延伸超越緊密連接屏障,並捕捉穿透角質層的外來抗原(見圖 14-10)。在此過程中,蘭格罕細胞樹突與 SG2 層角質細胞之間形成新的緊密連接,使外來抗原得以攝取而不擾亂緊密連接的屏障功能。蘭格罕細胞從表皮遷出,將捕捉到的抗原呈現給引流淋巴結 (draining lymph nodes) 中的 T 細胞,並被認為能誘導對所捕捉抗原的體液免疫 (humoral immunity)。¹⁰¹,¹⁰⁶
γ-δ T 細胞是 T 細胞的一個次要亞群,表現 T 細胞受體 γ 與 δ。在人類表皮中,γ-δ T 細胞罕見;然而,在小鼠表皮中,很大比例的 T 細胞為 γ-δ T 細胞。¹⁰⁷⁻¹¹⁰ 在小鼠表皮中,γ-δ T 細胞的樹突已被證明與帶有緊密連接的細胞的基底側對接 (dock),形成免疫突觸 (immunologic synapses),使 T 細胞的投射在角質細胞處極化並錨定。¹¹¹
維持皮膚屏障恆定的角質細胞形狀 (SHAPE OF KERATINOCYTES FOR SKIN BARRIER HOMEOSTASIS)
角質細胞依照規則的堆疊結構形成,相鄰細胞柱之間具有規則的鋸齒狀指狀交錯 (zigzag interdigitation) 模式(見圖 14-5B)。此種規則的指狀交錯堆疊結構在囓齒類耳部的皮膚中最為明顯,但也見於囓齒類其他身體部位的皮膚以及人類皮膚。¹¹² 對小鼠耳部皮膚表面角質細胞的經典掃描電子顯微鏡研究,以及近期對小鼠耳部帶有緊密連接的顆粒層細胞的免疫螢光顯微鏡研究揭示,細胞的基本形狀是角質層與顆粒層中克耳文十四面體(見表 14-1)的扁平變體⁴,¹¹³,¹¹⁴(圖 14-11)。角質細胞規則的指狀交錯模式被認為源自扁平化十四面體細胞規則的指狀交錯堆疊(見圖 14-5B 與 14-11C)。計算模擬與活體影像研究顯示,此規則結構是在顆粒層由來自棘層的細胞三維形狀的轉變所形成,儘管細胞形狀改變背後的精確分子機轉仍屬未知。⁴,¹¹⁵⁻¹¹⁷ 僅有單層細胞(SG2 層細胞)在側向細胞–細胞接觸的頂端邊緣形成緊密連接。因此,緊密連接形成於十四面體細胞的邊緣(見圖 14-11)。當一個形成緊密連接的細胞(舊細胞)被位於舊細胞正下方的新細胞所取代時,新細胞首先在舊細胞下方形成一個新的緊密連接多邊形 (tight junction polygon)。因此,在細胞替換過程中,每個緊密連接多邊形暫時變為雙邊 (double-edged)(見圖 14-11)。在此之後,舊細胞的緊密連接多邊形消失,最終舊細胞被擠出至緊密連接屏障之外(形成緊密連接屏障的 SG2 細胞層之外)。在每個細胞更新部位暫時形成的雙邊緊密連接多邊形維持了緊密連接屏障的恆定。⁴ 扁平化十四面體細胞規則的指狀交錯堆疊,不僅使雙邊緊密連接多邊形的暫時形成成為可能,也使細胞之間的空間關係以及橋粒細胞–細胞黏附在細胞更新期間得以維持,這可能負責表皮的物理強度。⁴ 將緊密連接形成活動限制於表皮 SG2 細胞層的分子機轉、在維持緊密連接屏障恆定的同時細胞更新的時空調控,以及十四面體細胞形狀產生的方式,目前皆屬未知。
抗菌屏障 (ANTIMICROBIAL BARRIER)
皮膚表面的抗菌特性 (ANTIMICROBIAL PROPERTY OF THE SKIN SURFACE)
皮膚持續遭遇微生物病原體。控制皮膚微生物群對於預防細菌與黴菌感染很重要。表皮細胞透過脫屑持續更新,禁止微生物在皮膚上的定植。皮膚表面本身的物理特性即可防止細菌生長,亦即低碳水化合物與低水分含量,以及弱酸性 pH 值(pH 為 5.6 至 6.4)。¹¹⁸⁻¹²⁰ 此弱酸性 pH 值是由各種物質所致,例如由皮脂腺分泌或源自角質層中磷脂質水解的游離脂肪酸、由外分泌腺 (eccrine glands) 分泌的乳酸、大多源自絲聚蛋白降解產物的尿刊酸,以及由微生物所產生的代謝物。¹²¹ 表皮也配備有一個由抗菌蛋白所組成的主動抗菌防禦系統(表 14-3)。
皮膚表面的抗菌蛋白 (ANTIMICROBIAL PROTEINS ON THE SKIN SURFACE)
抗菌蛋白是在演化上古老的先天免疫效應分子 (innate immune effectors),幾乎由所有植物與動物所產生。¹²²,¹²³ 抗菌蛋白對革蘭氏陽性與陰性細菌兩者都顯示廣泛的抗菌活性;有些甚至顯示抗黴菌或抗病毒活性。多數蛋白質的抗菌活性源自其獨特的結構特徵,使其能破壞微生物的膜,同時保持宿主細胞膜完整。這些蛋白質也作為「警報素」(alarmins),警示宿主細胞對損傷與微生物入侵作出反應。¹²⁴
許多蛋白質在皮膚中作為抗菌蛋白/警報素,包括短蛋白質,例如 β-防禦素 (β-defensins)、抗菌肽素 (cathelicidins)、皮膚抑菌素 (dermcidin)、銀屑病素 (psoriasin)、神經肽 (neuropeptides) 與趨化激素 (chemokines),以及較大的蛋白質,例如溶菌酶 (lysozymes)、彈性蛋白酶 (elastase)、補體 (complement) 與 S100 蛋白。這些抗菌蛋白的多樣性可能反映了皮膚表面微生物挑戰的複雜性與悠久歷史。襯於腸道、呼吸道、生殖道與皮膚表面的上皮細胞是抗菌蛋白的主要來源。每種上皮都有許多對各微環境所必需的特徵性抗菌蛋白。抗菌肽素與 β-防禦素是皮膚的兩種主要抗菌蛋白,並在體內顯示廣譜的抗菌活性。¹²⁵
產生抗菌蛋白的細胞 (CELLS THAT PRODUCE ANTIMICROBIAL PROTEINS)
角質細胞產生各種抗菌蛋白以防禦皮膚(圖 14-12)。特別是毛囊中的角質細胞,組成性地以比毛囊間表皮 (interfollicular epidermis) 角質細胞更高的程度產生抗菌肽素與 β-防禦素,可能是因為毛囊的微環境比毛囊間表皮的微環境更容易促進細菌定植。皮膚的分泌細胞,例如外分泌腺、頂泌腺 (apocrine) 與皮脂腺中的細胞,分泌它們自己的抗菌蛋白與脂質,並促成皮膚表面的抗菌活性。在真皮中,肥大細胞 (mast cells) 產生大量的抗菌肽素,並將其儲存於細胞內顆粒中。在皮膚損傷後,肥大細胞分泌抗菌肽素以抵抗細菌與病毒感染。¹²⁶ 此外,共生細菌 (commensal bacteria) 產生它們自己的抗菌蛋白。例如,表皮葡萄球菌(Staphylococcus epidermidis),皮膚微生物群的優勢共生細菌,產生數種抗菌蛋白。¹²⁷,¹²⁸ 這些源自宿主細胞與微生物的各種抗菌蛋白,能調節或調控皮膚微生物群的組成。
抗菌蛋白的功能 (FUNCTIONS OF ANTIMICROBIAL PROTEINS)
抗菌蛋白具有各種作用機轉¹²²,¹²⁹,¹³⁰;許多以微生物的細胞壁或細胞膜結構為標的。例如,防禦素與抗菌肽素通常帶正電(cationic),並透過靜電交互作用與細菌膜表面交互作用。某些防禦素在細菌膜上形成孔洞以破壞膜的完整性並促進細菌溶解。抗菌肽素與細菌膜結合並促進膜的插入與破壞。數種抗菌蛋白的作用機轉仍不清楚。越來越多的證據顯示,某些抗菌蛋白透過趨化激素受體與類鐸受體調節免疫訊號傳遞。抗菌蛋白不僅刺激各種細胞類型分泌趨化激素與細胞激素,也利用趨化活性 (chemotactic activity) 以直接或間接的機轉招募白血球 (leukocytes),以修飾發炎反應。¹²²,¹³¹⁻¹³³ 不同類型抗菌蛋白的趨化活性彼此不同,各招募不同類型的細胞。例如,LL-37,抗菌肽素家族抗菌肽的人類成員,招募嗜中性球 (neutrophils)、T 細胞、單核球 (monocytes) 與肥大細胞。LL-37 也透過粒線體抗病毒訊號傳遞途徑 (mitochondrial antiviral signaling pathway) 誘導角質細胞,以促進干擾素-β(interferon-β)的生成與分泌,從而活化抗病毒反應。¹³⁴ 類鐸受體是用於模式辨識的跨膜受體,並由微生物分子的保守結構模式(例如脂多糖 [lipopolysaccharides]、鞭毛 [flagella]、內毒素 [endotoxins]、RNA 與 DNA)所活化。¹³⁵ 類鐸受體的訊號傳遞不限於此類微生物分子,也可由某些抗菌蛋白所啟動,使免疫細胞反應得以被修飾。¹³⁶
新生兒皮膚的表皮屏障 (EPIDERMAL BARRIER OF NEWBORN SKIN)
足月新生兒在出生時即具有發育完善且具功能的皮膚屏障。¹³⁷ 雖然與成人皮膚相比,嬰兒皮膚的角質層較薄、天然保濕因子濃度較低,且經皮水分散失較大,但健康足月新生兒出生時的經皮水分散失等於或低於成人皮膚(表 14-4)。¹³⁸⁻¹⁴⁰
角質層在子宮內於暴露於羊水 (amniotic fluid) 的情況下發育,然而廣泛的水分暴露卻會破壞成人皮膚的角質層屏障。子宮內角質層屏障發育的一個假定機轉,是胎兒生物膜胎脂 (vernix caseosa) 的形成,胎脂在第三(最後)孕期 (trimester) 覆蓋整個皮膚。¹⁴¹ 胎脂是水、蛋白質與脂質的複雜混合物;充滿水分而腫脹的胎兒角質細胞分布於脂質的無定形混合物中。胎脂保護表皮免於羊水侵害,並促進其下方角質層的形成。據報告,與出生後立即去除胎脂相比,出生後保留胎脂在出生後 24 小時時會帶來更高的皮膚水合 (skin hydration) 與更低的皮膚表面 pH 值。¹⁴¹ 此外,胎脂含有抗菌劑,包括溶菌酶與乳鐵蛋白 (lactoferrin),並展現抗黴菌與抗細菌活性。¹⁴²⁻¹⁴⁴ 此證據暗示,保留胎脂而非如世界衛生組織(World Health Organization)所建議的在出生後立即去除,具有其價值。¹⁴⁵
與足月嬰兒不同,早產兒在出生時具有脆弱的表皮屏障與機械性脆弱的真皮。妊娠 28 週前出生的嬰兒缺乏胎脂的覆蓋。極低出生體重 (very low birth weight) 的新生兒由於皮膚屏障不足,有較高的皮膚損傷風險。妊娠 25 週分娩的嬰兒在出生時僅有少數幾層角質層,並顯示約 70 g/m²/hr 的經皮水分散失。¹⁴⁶ 早產新生兒出生時的經皮水分散失逐漸降低至約 7 g/m²/hr,直到妊娠 35 週左右,此時表皮屏障在子宮內已發育完善。表皮屏障在出生後持續發育,但即使在分娩後 1 個月,極早產兒 (extremely preterm infants) 的經皮水分散失仍顯著高於足月兒。¹⁴⁶
對抗物理性應力的屏障 (BARRIER AGAINST PHYSICAL STRESSES)
對抗機械性應力的物理結構 (PHYSICAL STRUCTURE PROTECTING AGAINST MECHANICAL STRESSES)
保護我們身體免於各種機械性物理刺激的皮膚,由堅硬與柔軟組織的條帶 (stripes) 所組成,亦即堅硬的角質層、柔軟的角質細胞層、堅硬的真皮膠原組織,以及柔軟的下皮 (hypodermis,富含脂肪細胞 adipocytes) 緩衝墊。各條帶的物理強度缺陷以及邊界缺陷,會造成各種導致物理脆弱性的疾病,例如表皮鬆解性魚鱗癬 (epidermolytic ichthyosis);單純型、交界型與失養型表皮鬆解水疱症 (simple, junctional, and dystrophic epidermolysis bullosa);以及埃勒斯-當洛斯症候群 (Ehlers-Danlos syndromes)(見第 15、47、60 與 72 章)。
對抗紫外線應力的屏障 (BARRIERS AGAINST ULTRAVIOLET STRESSES)
空氣–皮膚界面的反射、反式尿刊酸的吸收,以及經由平行於皮膚表面排列的角蛋白絲的繞射 (diffraction),限制了紫外線輻射滲透進入角質層。在表皮具活性的細胞層中,黑色素是吸收紫外線照射並保護基因組 DNA 免於紫外線誘導損傷的主要因子。雖然表皮上層細胞由於其在表皮中的位置,可能比基底層細胞遭受更大的紫外線照射 DNA 損傷,但這些細胞被緊密控制於非增生狀態,並經由細胞更新持續從表皮中被清除,從而禁止腫瘤生成 (tumorigenesis)。在具增生能力的基底層細胞中,由紫外線照射誘導的基因組 DNA 損傷立即被 DNA 修復酶所修復(見第 130 章)。
出汗以對抗熱應力 (SWEATING TO PROTECT AGAINST HEAT STRESSES)
皮膚是對抗熱與冷應力的保護器官,並在控制體溫上扮演重要角色。在人類中,冷卻主要透過從外分泌汗腺 (eccrine sweat glands) 分泌的水分蒸發來達成。中暑 (Thermoplegia) 傾向發生於患有先天性與後天性無汗症 (anhidrosis) 或少汗症 (hypohidrosis) 的病人。人類皮膚在所有哺乳動物中具有報告中最高密度的外分泌汗腺。¹⁴⁷ 多數哺乳動物汗腺很少,但具有覆蓋其身體的濃密毛髮。由於分泌的汗液在皮毛下不易蒸發,多毛的哺乳動物不利用汗液冷卻其身體是合理的。相對地,人類無毛的皮膚具有高密度的外分泌汗腺,並能經由體表的水分蒸發進行有效的冷卻。有趣的是,在小鼠足墊 (footpad) 中,外分泌腺與毛囊的密度由一個轉錄因子 En1 反向決定。¹⁴⁸
對抗熱應力的脈管系統與血流控制 (VASCULATURE AND BLOOD FLOW CONTROL AGAINST HEAT STRESSES)
冷卻與維持熱量部分取決於皮膚表面血流的控制,以及動脈與靜脈之間的逆流熱交換 (countercurrent heat exchange)。¹⁴⁹,¹⁵⁰ 當體溫上升時,皮膚表面血管擴張,皮膚顯得發紅。血液經由汗液蒸發以及體表的輻射誘導冷卻 (radiation-induced cooling) 而冷卻下來(圖 14-13)。在寒冷的空氣中,皮膚因血管收縮以減少血流並經由輻射冷卻將熱量流失降至最低而轉為蒼白。因此,皮膚調節體表的血流以維持體溫的恆定。在皮膚中,動脈與靜脈緊密並列地走行。動脈與靜脈之間的血管逆流熱交換已被提出作為恆溫動物 (homeothermic animals) 在極寒或極熱條件下維持核心體溫的機轉。例如,鴨腿中傳入動脈 (afferent arteries) 與傳出靜脈 (efferent veins) 之間的逆流熱交換防止足墊的熱量流失,使鴨子能在冰冷的水中划水。在小鼠皮膚中,動脈與靜脈緊密並列地排列。在基因工程小鼠中,動靜脈排列的喪失導致體表更大的熱量流失,因為較溫暖的動脈血到達體表時未經由對抗從體表返回的靜脈血的逆流熱交換而預先冷卻。核心體溫因較冷的靜脈血返回身體核心而降低。¹⁵¹ 這些觀察,連同對人類動脈與靜脈血液溫度的測量¹⁵⁰,顯示人類在極端條件下也利用血管逆流熱交換來維持核心體溫(見圖 14-13)。
結論 (CONCLUSIONS)
直接暴露於外在環境的皮膚顯示出物種特異性的適應,導致脊椎動物與哺乳動物之間,甚至人類種族與人體各部位之間驚人的多樣性。皮膚科醫師了解人類皮膚如何適應外在環境,以及這些適應如何受疾病影響,至關重要。皮膚是一個由外胚層 (ectoderm)、中胚層 (mesoderm) 與皮膚微生物群所組成的複雜超級器官 (superorgan)。對人類皮膚、其微生物群,以及皮膚所暴露的外在環境進行整合性的理解,對於進一步研究正常皮膚功能以及各種皮膚疾病的分子致病機轉是必要的;這將最終改善人類健康。
致謝 (ACKNOWLEDGMENTS)
作者感謝 Ehrhardt Proksch 與 Jens-Michael Jensen 的貢獻,兩位是 Fitzpatrick’s Dermatology in General Medicine 第 8 版第 47 章「皮膚作為保護器官(Skin as an Organ of Protection)」的作者。
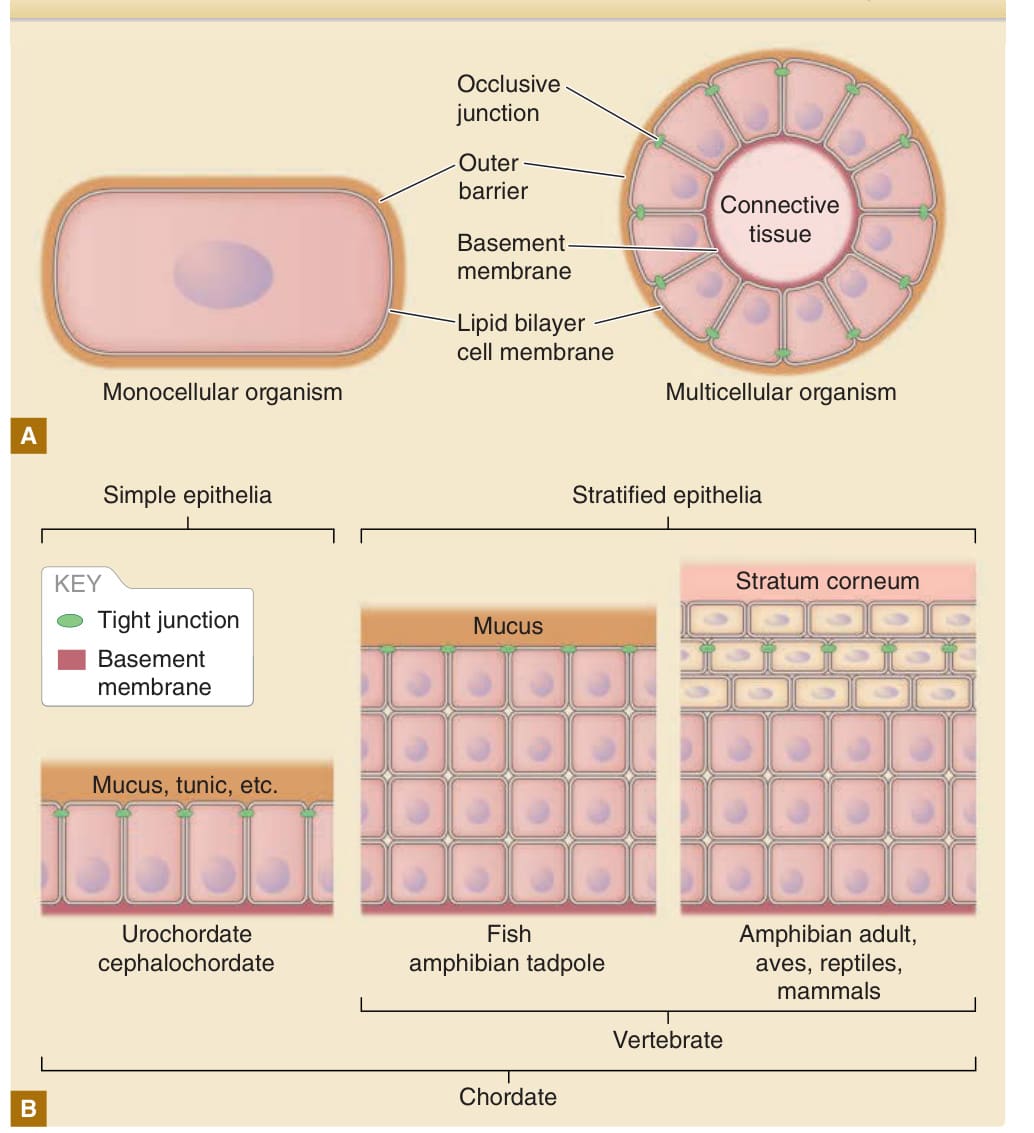
圖 14-1:單細胞與多細胞生物表面屏障的基本結構。A,脂質雙層細胞膜 (lipid bilayer cell membrane) 是單細胞生物與多細胞生物體表的基本擴散屏障。在多細胞生物中,細胞間隙由封閉性連接所封閉,以限制屏障經由細胞旁路徑滲漏(見圖 14-9)。本質上,單細胞生物與多細胞生物都配備有額外的外部屏障(即單細胞生物的細胞壁,以及多細胞生物的黏液、角質層 [cuticles]、被囊基質或角質層 [stratum corneum])。B,複層上皮 (stratified epithelium) 與角質層的發展是脊椎動物皮膚演化的兩大事件。在外部屏障之下,緊密連接封閉脊椎動物的細胞間隙。
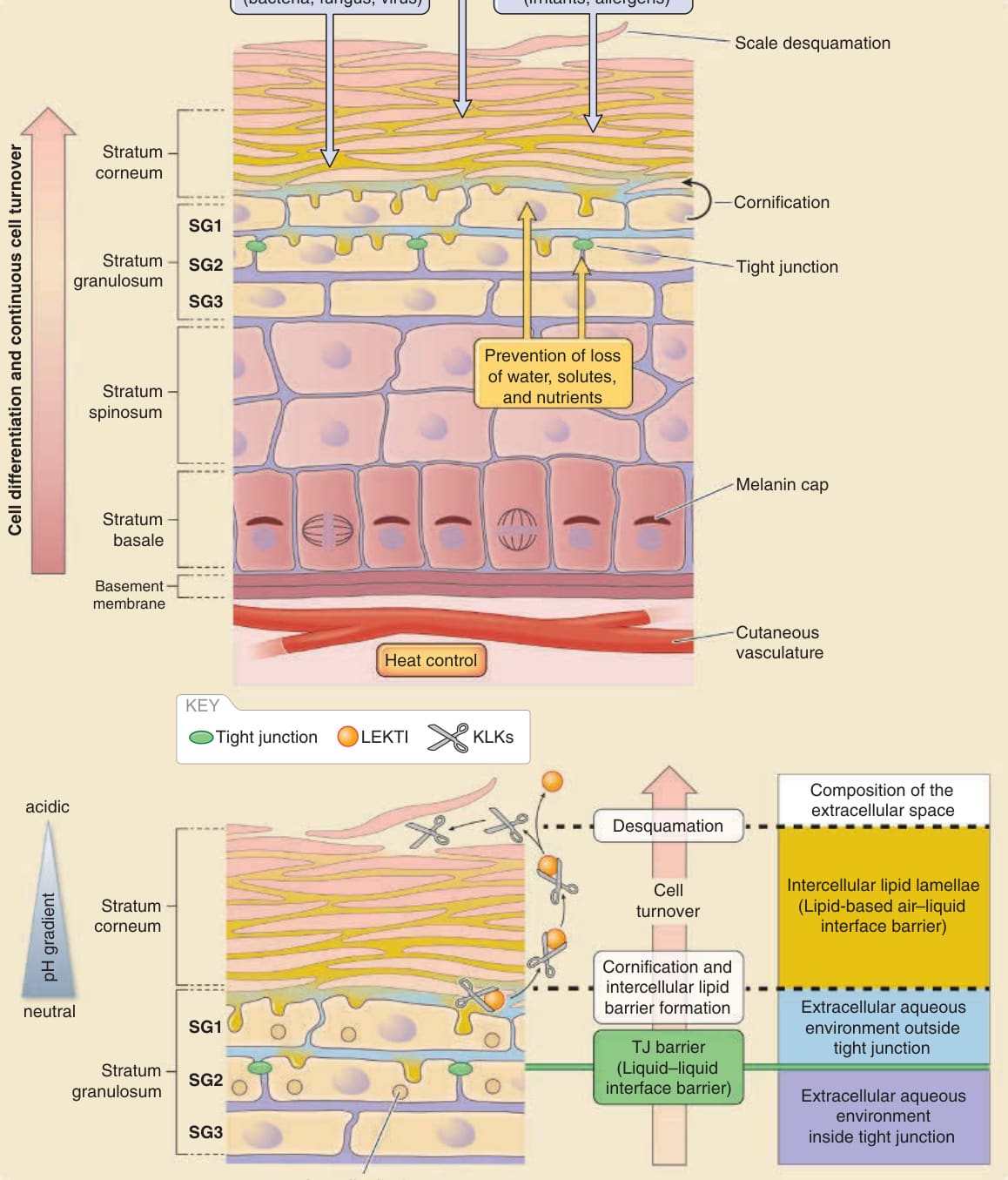
圖 14-2:表皮的基本結構與主要的表皮屏障。A,表皮屏障保護身體免於各種由外向內的物理、化學與微生物侵害,並防止水分與溶質由內向外滲漏。黑色素帽 (Melanin caps) 保護基底細胞的基因組 DNA 免於紫外線 (ultraviolet, UV) 損傷。細胞持續更新以更新表皮及其屏障。在角質層下方的顆粒層中存在至少三個細胞層(由外而內分別為 SG1、SG2 與 SG3),此處緊密連接 (tight junctions, TJs) 封閉 SG2 細胞之間的細胞間隙。B,細胞間隙在角質層中被脂質填滿,而在角質層下方則被水填滿。細胞外水環境被認為由緊密連接屏障分為兩部分(緊密連接屏障外側的細胞外空間,淺藍色;緊密連接屏障內側的細胞外空間,紫色)。SG1 細胞在緊密連接屏障外側準備角質化。層板小體 (lamellar bodies) 的內容物(脂質、抗菌蛋白、蛋白酶與蛋白酶抑制劑)基本上由 SG2 細胞的頂端表面胞吐至緊密連接屏障外側。激肽釋放酶蛋白酶(kallikrein proteases, KLKs)在角質層上層經由低 pH 誘導的淋巴上皮 Kazal 型相關抑制劑(lymphoepithelial Kazal-type-related inhibitor, LEKTI)的解離而活化,以進行脫屑。(修改自 Proksch E, Jensen J-M. Skin as an organ of protection. In: Goldsmith LA, Katz SI, Gilchrest BA, et al, eds. Fitzpatrick’s Dermatology in General Medicine, 8th ed. New York: McGraw-Hill; 2012.)
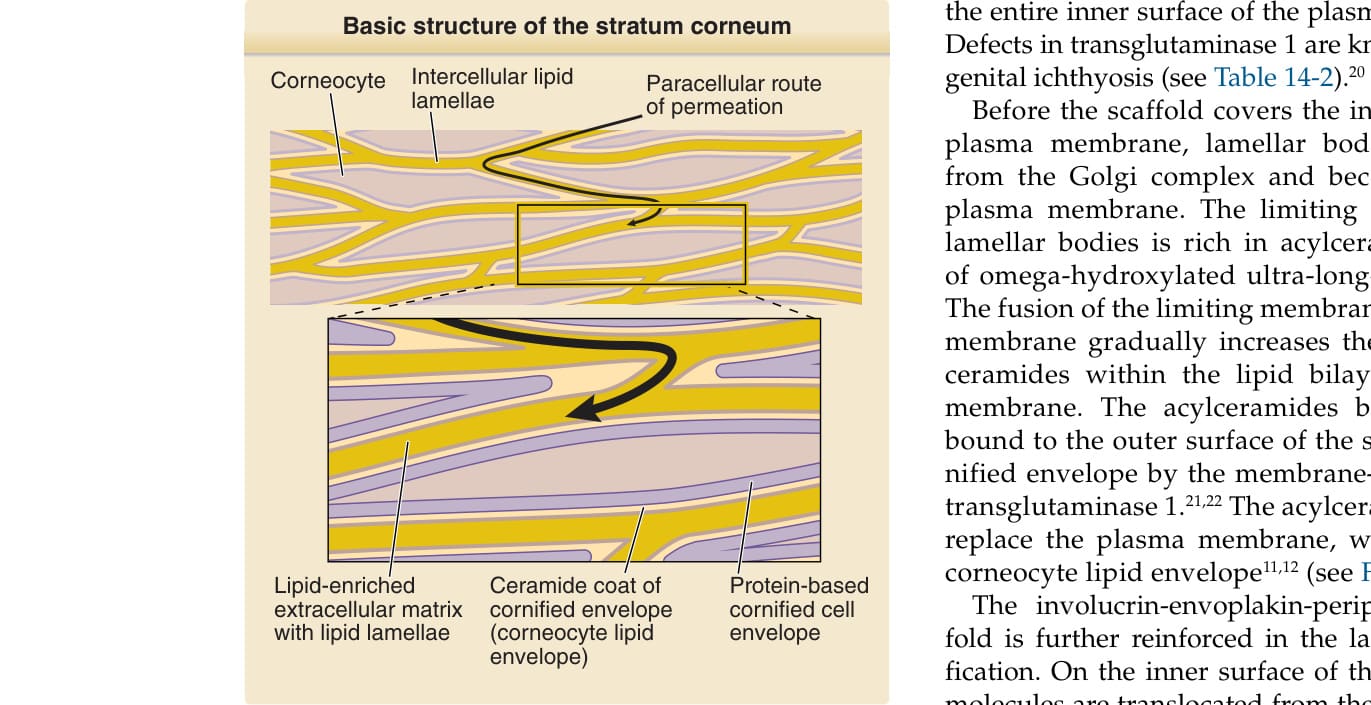
圖 14-3:角質層的基本結構。角質層由以蛋白質為基礎的磚塊(角質細胞 corneocytes)與以脂質為基礎的灰泥(細胞間脂質層板 intercellular lipid lamellae)所構成。角質細胞的表面被一層薄薄的神經醯胺(脂質包膜 lipid envelope)所覆蓋。此結構如何產生的細節顯示於圖 14-6。(修改自 Proksch E, Jensen J-M. Skin as an organ of protection. In: Goldsmith LA, Katz SI, Gilchrest BA, et al, eds. Fitzpatrick’s Dermatology in General Medicine, 8th ed. New York: McGraw-Hill; 2012.)
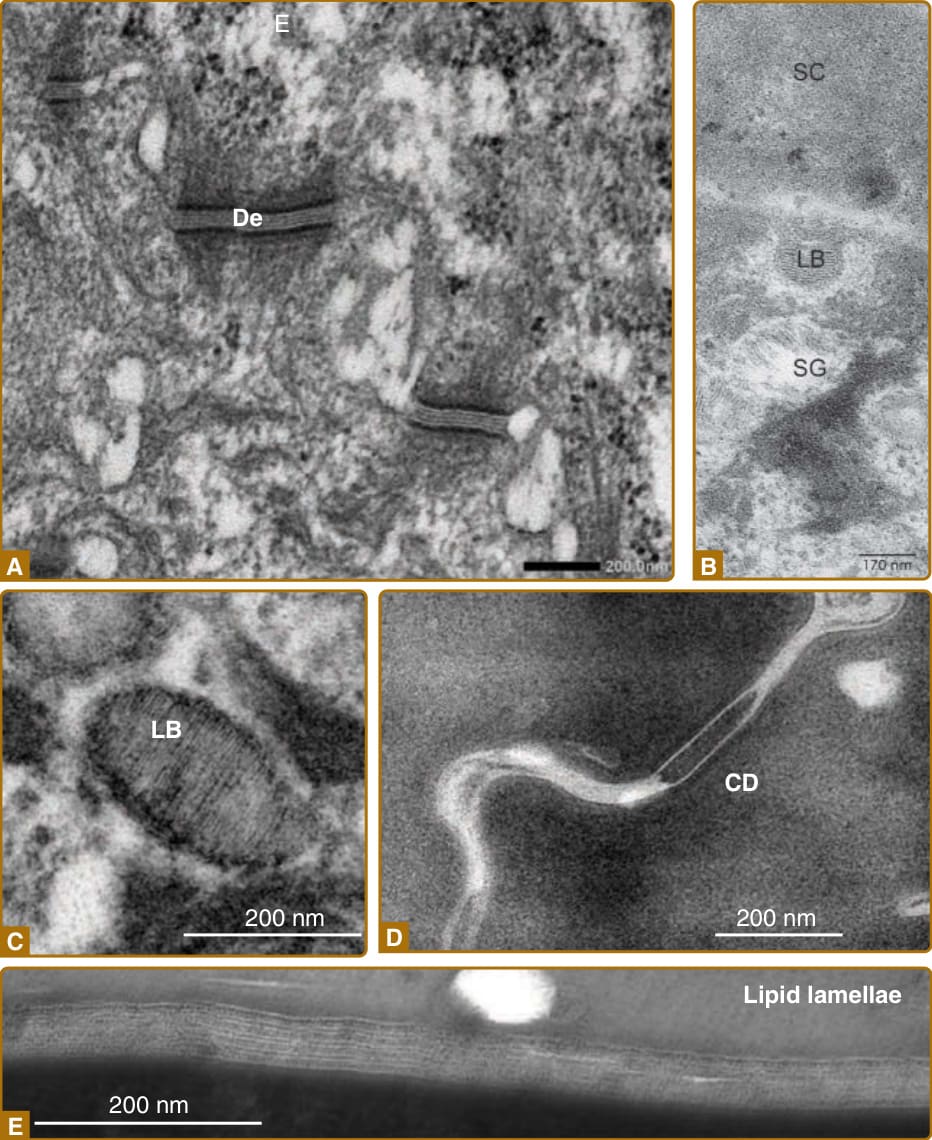
圖 14-4:與角質化相關的主要結構,於電子顯微鏡下所見。A,黏附於棘層細胞的橋粒(desmosomes, De)。B,一個層板小體(lamellar body, LB)於 SG1 細胞頂端表面將其內容物胞吐至細胞外空間。C,層板小體(LB)的內容物顯示脂質的條紋。D,角質橋粒(corneodesmosomes, CD),角質細胞之間的細胞–細胞黏附複合體。E,角質細胞之間的細胞間脂質層板。(影像 C 與 E,經 Akemi Ishida-Yamamoto 醫師許可使用。影像 A 與 D,經 Toshihiro Nagai 醫師許可使用。影像 B 修改自 Proksch E, Jensen J-M. Skin as an organ of protection. In: Goldsmith LA, Katz SI, Gilchrest BA, et al, eds. Fitzpatrick’s Dermatology in General Medicine, 8th ed. New York: McGraw-Hill; 2012.)
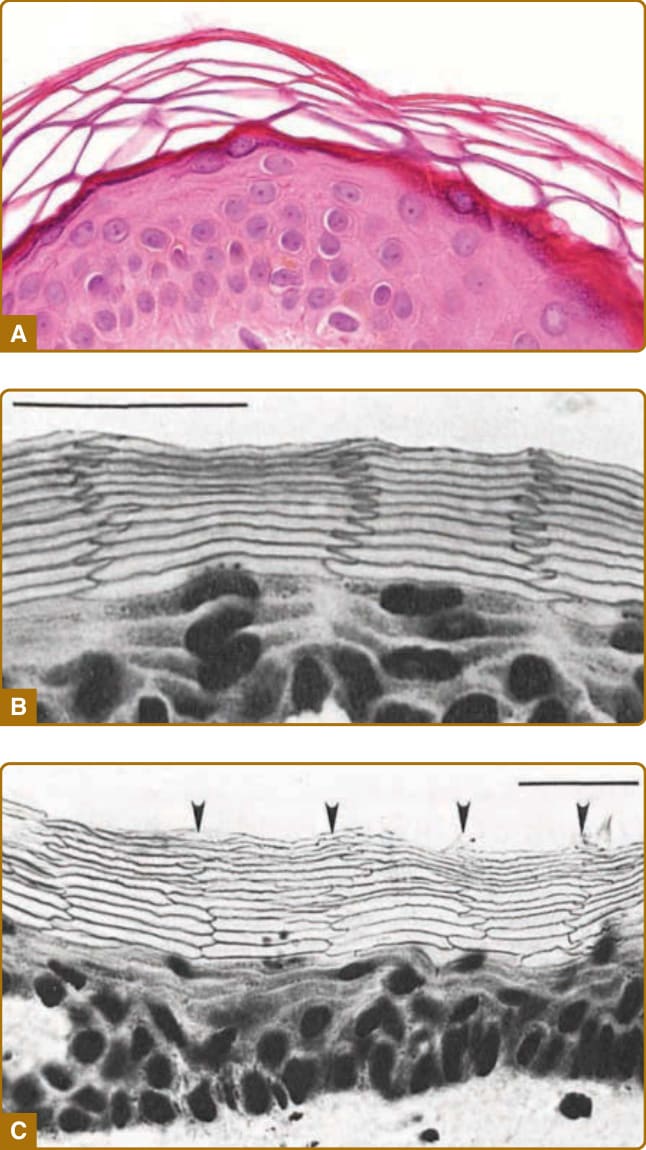
圖 14-5:橫切面下表皮的形態。A,人類皮膚石蠟包埋切片的蘇木精與伊紅染色,顯示角質層的籃網狀模式 (basket weave–like pattern)。B,倉鼠耳部皮膚的冷凍切片,以亞甲基藍 (methylene blue) 染色,並於 pH 12 的鹼性溶液中膨脹,顯示角質細胞的規則排列。可見扁平顆粒層細胞與無核角質細胞的顯著有序結構。角質細胞顯示細胞柱之間規則的鋸齒狀指狀交錯模式。C,人類腹部皮膚的冷凍切片,以亞甲基藍染色並於鹼性溶液中膨脹,顯示角質細胞部分規則的排列。(影像 B 取自 Mackenzie IC. Ordered structure of the epidermis. J Invest Dermatol. 1975;65:45-51,經許可使用;影像 C 取自 Mackenzie IC, Zimmerman K, Peterson L. The pattern of cellular organization of human epidermis. J Invest Dermatol. 1981;76:459-461,經許可使用。Copyright © The Society for Investigative Dermatology.)
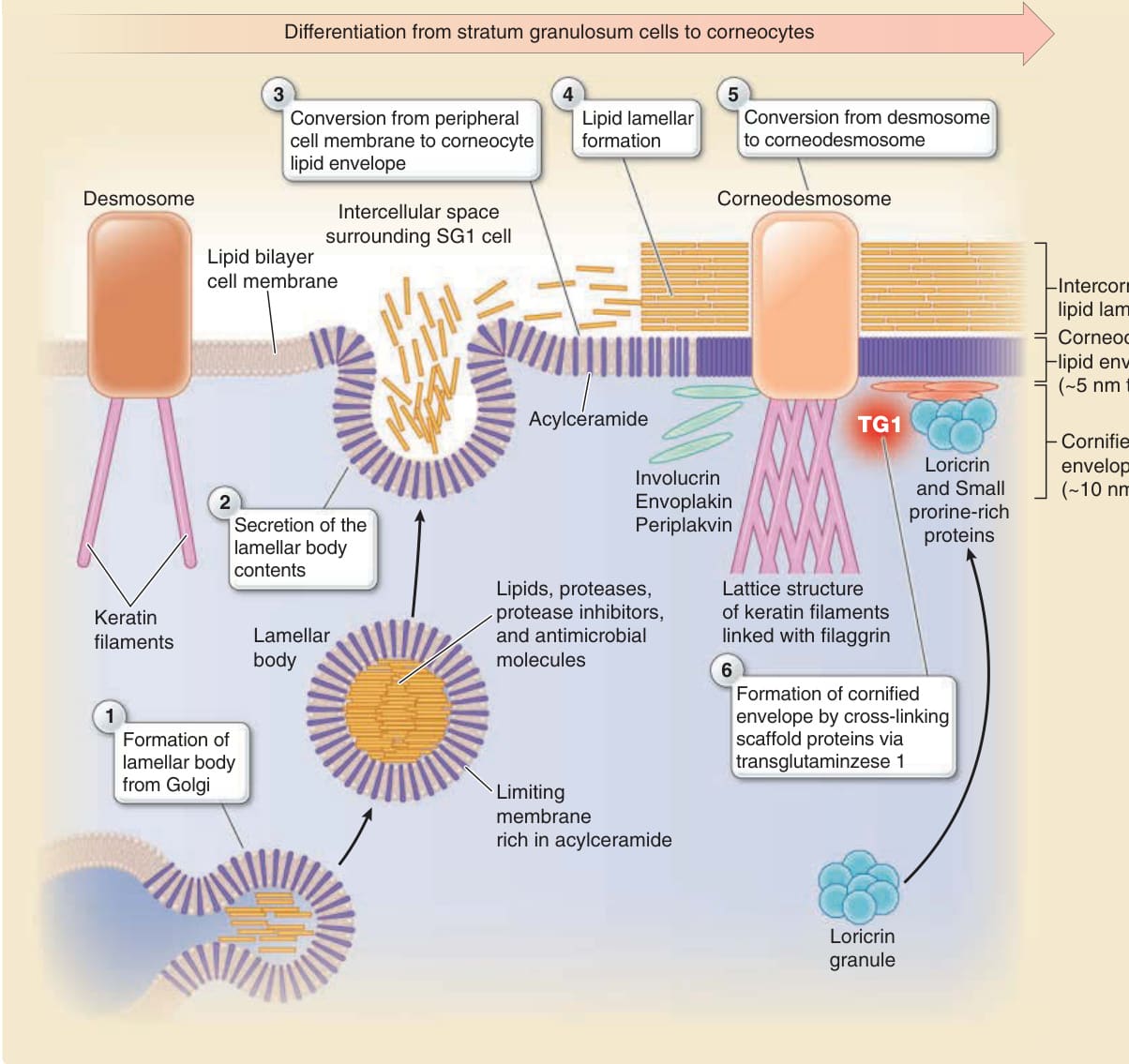
圖 14-6:角質化細胞包膜、角質細胞脂質包膜與角質細胞間脂質層板的形成。角質化的時間依賴性變化(從 SG2 細胞經由 SG1 細胞到角質細胞)由左至右顯示。一層角質化細胞包膜在脂質雙層細胞膜之下,經由兜膜蛋白 (envoplakin)、外周斑蛋白 (periplakin) 與兜甲蛋白 (involucrin) 被轉麩醯胺酶 1(transglutaminase 1, TG1)交聯而形成,並在角質化後期階段由兜甲蛋白 (loricrin) 進一步強化。層板小體將其內容物胞吐至緊密連接外側的細胞外空間。層板小體的界膜含有大量的醯基神經醯胺,取代脂質雙層細胞膜以形成角質細胞脂質包膜。結果,角質化細胞包膜被角質細胞脂質包膜所覆蓋。從層板小體胞吐的脂質在角質細胞脂質包膜上形成角質細胞間脂質層板,以填滿細胞間隙並形成角質層的抗水屏障。從層板小體胞吐的角質橋粒素整合進橋粒,橋粒轉化為角質橋粒。層板小體也胞吐蛋白酶與蛋白酶抑制劑以控制脫屑,以及抗菌蛋白(見圖 14-12)。
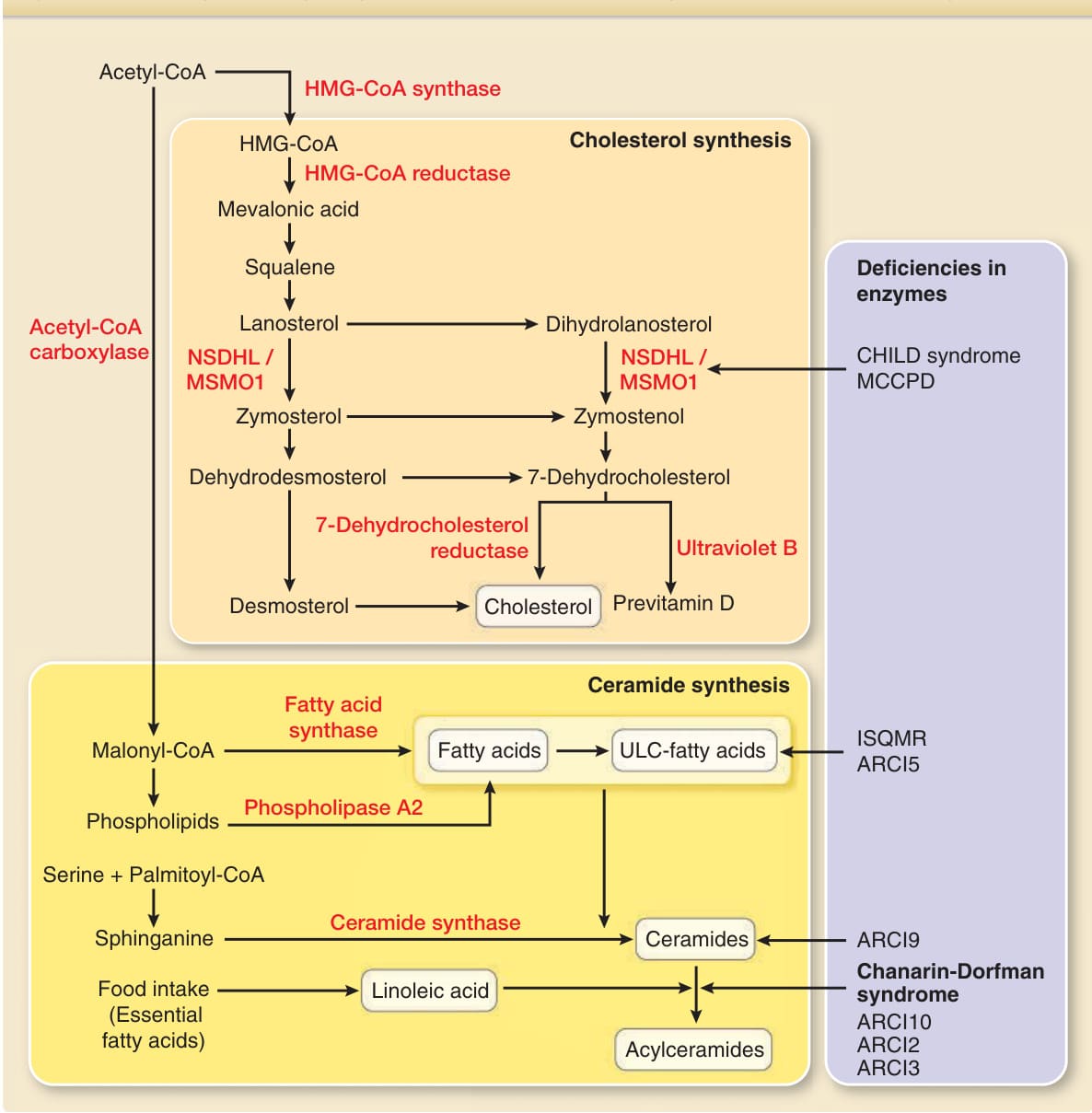
圖 14-7:膽固醇、游離脂肪酸、神經醯胺與醯基神經醯胺的合成途徑、關鍵酵素,以及酵素缺陷。CoA,輔酶 A (coenzyme A);HMG-CoA,羥甲基戊二醯輔酶 A (hydroxymethylglutaryl CoA);CHILD syndrome,伴有魚鱗癬樣紅皮症與肢體缺陷的先天性偏側發育不全症候群 (congenital hemidysplasia with ichthyosiform erythroderma and limb defects syndrome);MCCPD,小頭症、先天性白內障與銀屑病樣皮膚炎 (microcephaly, congenital cataract, and psoriasiform dermatitis);ISQMR,魚鱗癬、痙攣性四肢癱瘓與智能遲緩 (ichthyosis, spastic quadriplegia, and mental retardation);ARCI,體染色體隱性遺傳先天性魚鱗癬 (autosomal recessive congenital ichthyosis);NSDHL,NAD(P) 依賴性類固醇去氫酶樣 (NAD(P) dependent steroid dehydrogenase-like);MSMO1,甲基固醇單加氧酶 1 (methylsterol monooxygenase 1);ULC,超長鏈 (ultra-long-chain)。
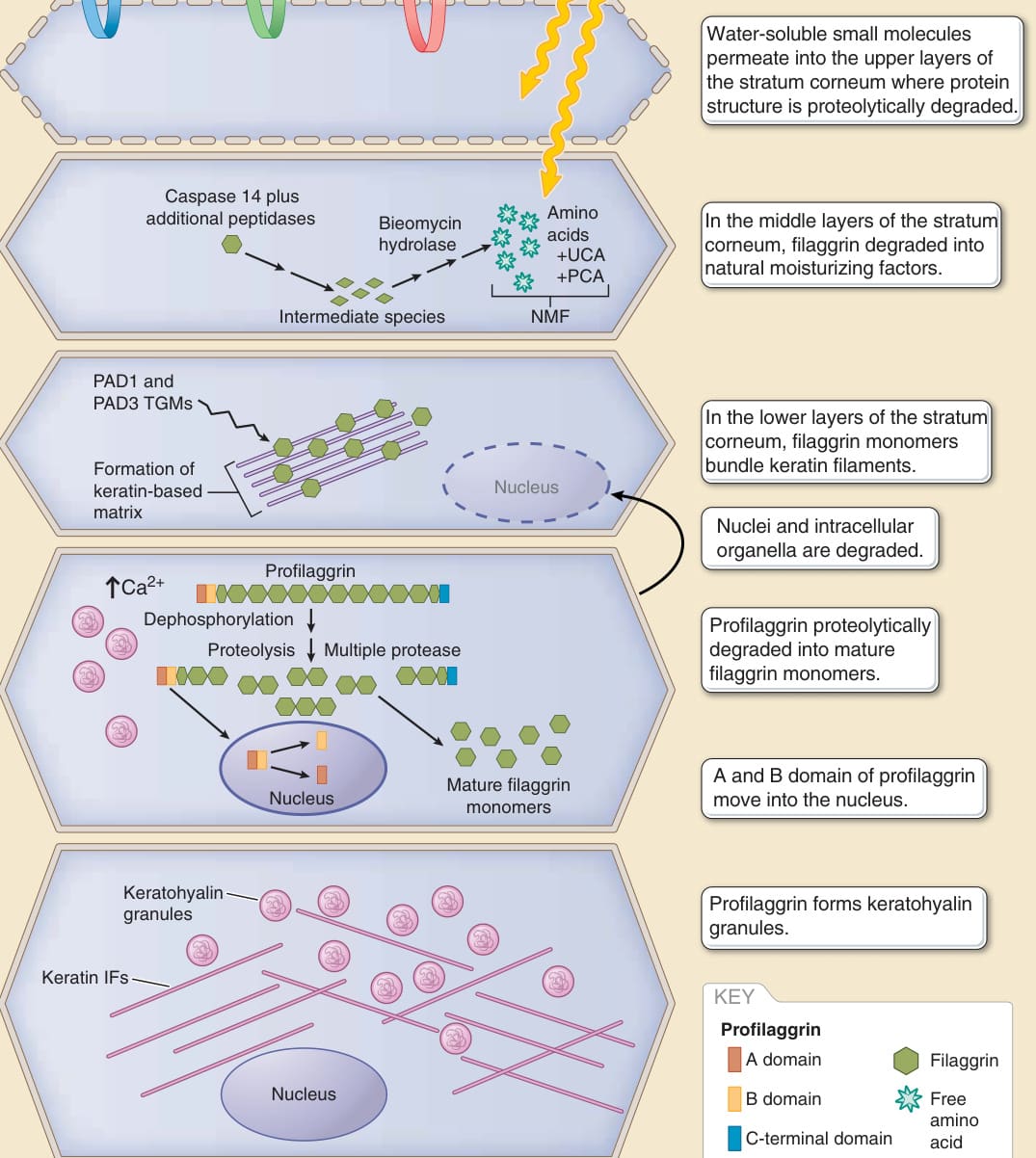
圖 14-8:角質層的多區域結構與絲聚蛋白的多步驟成熟與降解。角質層被認為由至少三種經歷不同分化步驟的角質細胞所組成(角質層下層、中層與上層的角質細胞)。NMF,天然保濕因子 (natural moisturizing factor);PAD,肽基精胺酸去亞胺酶 (peptidylarginine deiminase);PCA,2-吡咯啶酮-5-羧酸 (2-pyrrolidone-5-carboxylic acid);IF,中間絲 (intermediate filament);SC,角質層 (stratum corneum);TGM,轉麩醯胺酶 (transglutaminase);UCA,尿刊酸 (urocanic acid)。(修改自 McAleer MA, Irvine AD. The multifunctional role of filaggrin in allergic skin disease. J Allergy Clin Immunol. 2013;131(2):280-291.)
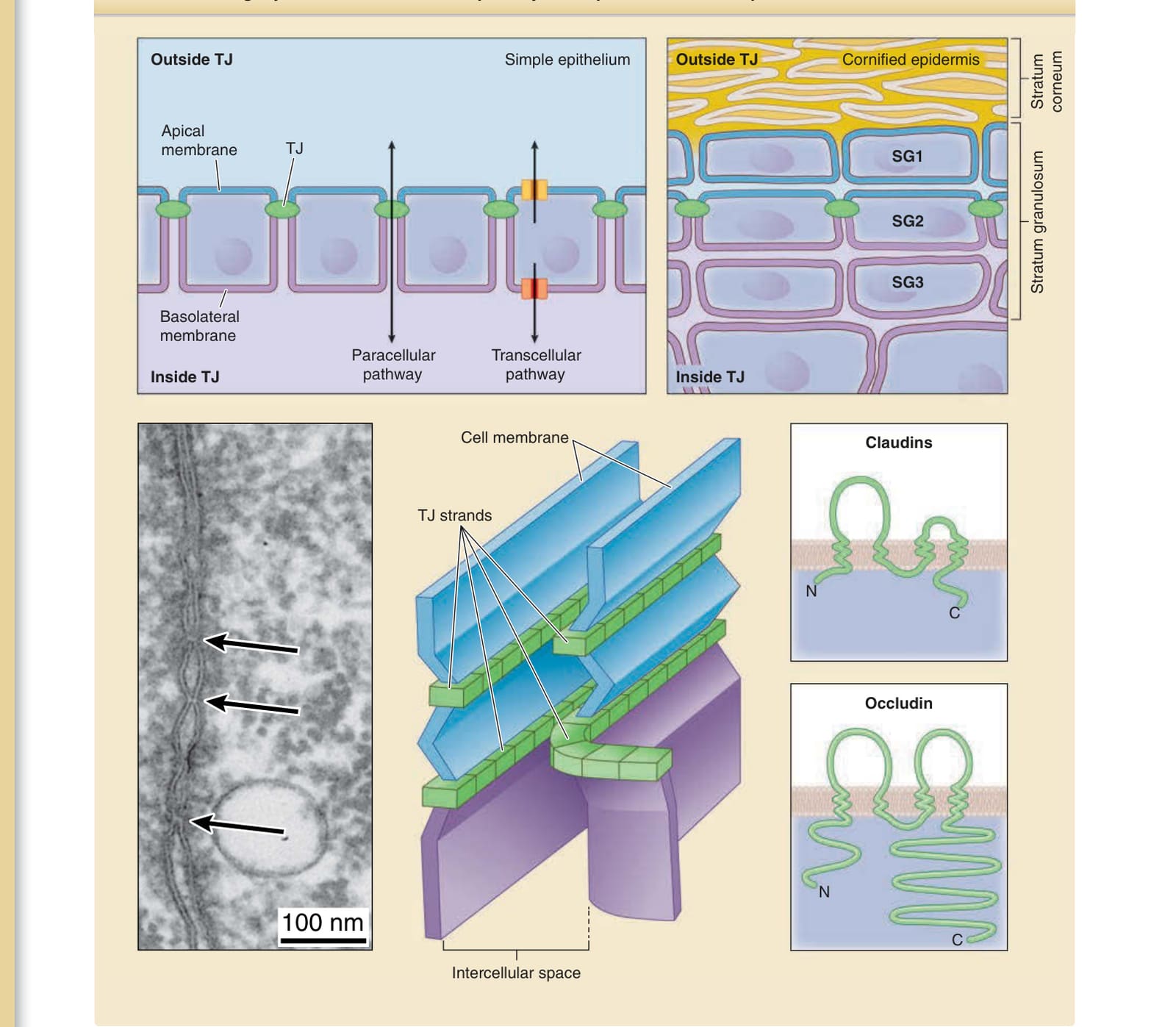
圖 14-9:單層與複層上皮細胞片中的緊密連接(tight junction, TJ)屏障。在單層上皮中,細胞外環境與質膜在緊密連接屏障處被分為兩部分。溶質經由細胞旁與細胞穿透途徑在兩個隔室之間移動。在表皮中,越來越多的證據顯示,細胞外環境與質膜也在 SG2 細胞之間形成的緊密連接屏障處被分為兩部分,即緊密連接屏障外側與內側的細胞外環境,以及頂端與基底側細胞膜。在電子顯微照片中(左圖),緊密連接呈現為一個「親吻點」(kissing point,箭頭),即兩個質膜彼此相對之處。示意圖顯示細胞膜上緊密連接索的結構以及緊密連接的主要跨膜蛋白。C,C 端 (C-terminus);N,N 端 (N-terminus)。(影像經 Hiroyuki Sasaki 醫師許可使用。)
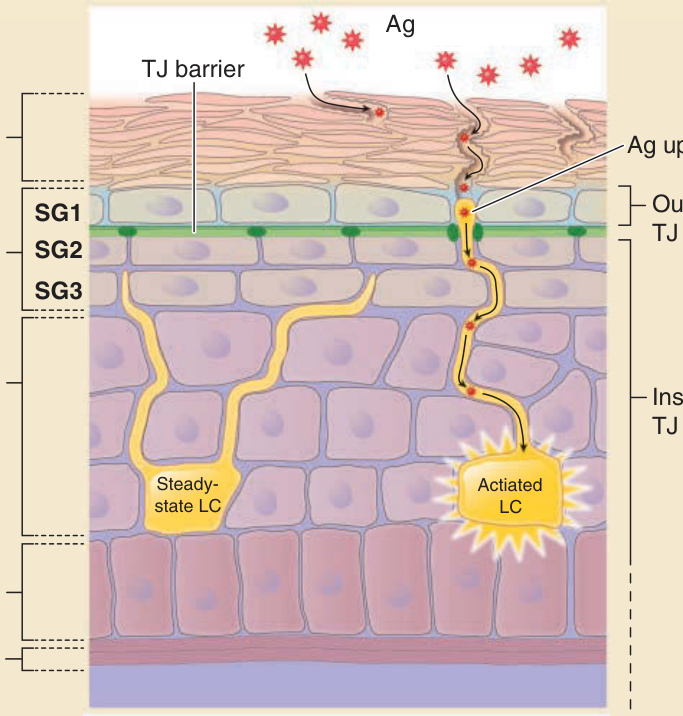
圖 14-10:角質層、緊密連接(tight junctions, TJs)與蘭格罕細胞的空間位置。SG2 細胞形成緊密連接(深綠色實心圓)。SG1 細胞位於緊密連接屏障外側(淺藍色)。相對地,SG3 細胞、棘層細胞與蘭格罕細胞位於緊密連接屏障內側(紫色)。穩定狀態下蘭格罕細胞的樹突(左側黃色細胞)位於緊密連接之下,但在其活化狀態下(右側黃色細胞)穿透緊密連接屏障以進入緊密連接屏障外側。抗原(antigen, Ag)的攝取發生於緊密連接屏障外側的樹突末端(以 Ag uptake 標示)。
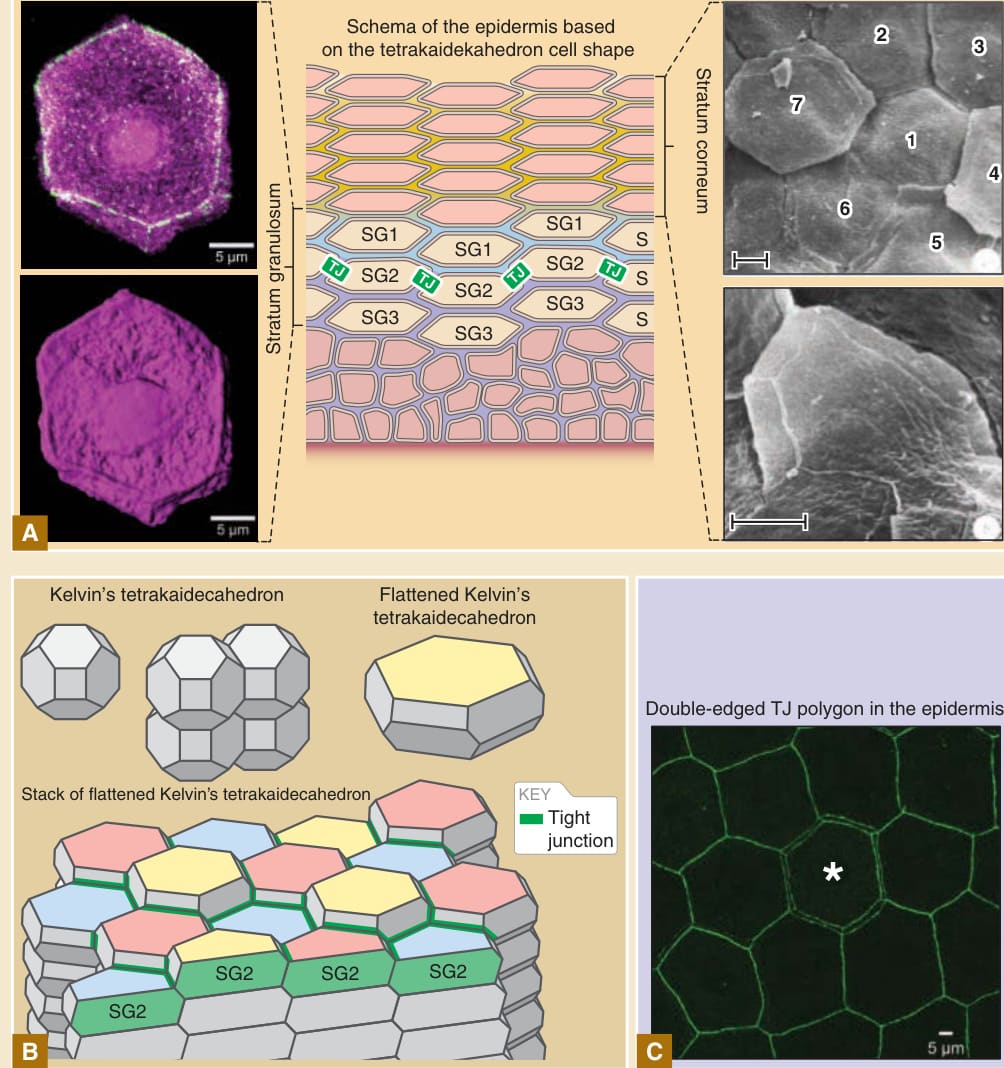
圖 14-11:角質細胞與形成緊密連接(tight junction, TJ)細胞的三維細胞形狀。A,免疫螢光顯微鏡視野下形成緊密連接細胞的三維多面體形狀(左圖),以及掃描電子顯微鏡視野下角質層表面角質細胞的等效形狀(右圖),顯示顆粒層細胞與角質細胞的基本形狀是克耳文十四面體 (Kelvin’s tetrakaidecahedron) 的扁平變體。B,克耳文十四面體、克耳文十四面體的扁平變體,以及扁平化克耳文十四面體細胞的規則指狀交錯堆疊。SG2 細胞顯示於細胞柱的頂端。SG2 細胞的三個不同 Z 軸位置以黃色、粉紅色與藍色顯示於細胞的頂端表面。緊密連接(綠色邊緣)位於 SG2 細胞間細胞–細胞接觸的頂端邊緣。C,小鼠耳部皮膚中緊密連接(以抗 ZO-1 抗體染色的綠色邊緣)的正面視圖 (en face view)。本質上,緊密連接多邊形為單邊。雙邊多邊形(星號)是形成新緊密連接多邊形以將細胞從緊密連接屏障內側重新定位至外側的部位。D,在細胞從緊密連接屏障內側向外側更新期間,雙邊緊密連接多邊形(星號)形成的概念模型。一個新的緊密連接多邊形在中央黃色細胞下方形成(星號,左圖),導致雙邊多邊形的形成(中圖)。然後,外側的舊多邊形被降解(右圖)。左圖中央的黃色細胞(星號)在雙邊多邊形暫時形成(中圖)後,從緊密連接屏障內側(左圖)重新定位至緊密連接屏障外側(右圖)。(影像 A 右側的掃描電子顯微鏡圖片,經 Springer Nature 許可複製自 Allen TD, Potten CS. Significance of cell shape in tissue architecture. Nature. 1976;264:545-547。)A 與 C 部分的免疫螢光圖片以及 B 與 D 中的示意圖取自 Yokouchi M, Atsugi T, Logtestijn M, et al. Epidermal cell turnover across tight junctions based on Kelvin’s tetrakaidecahedron cell shape. Elife. 2016;5:e19593.
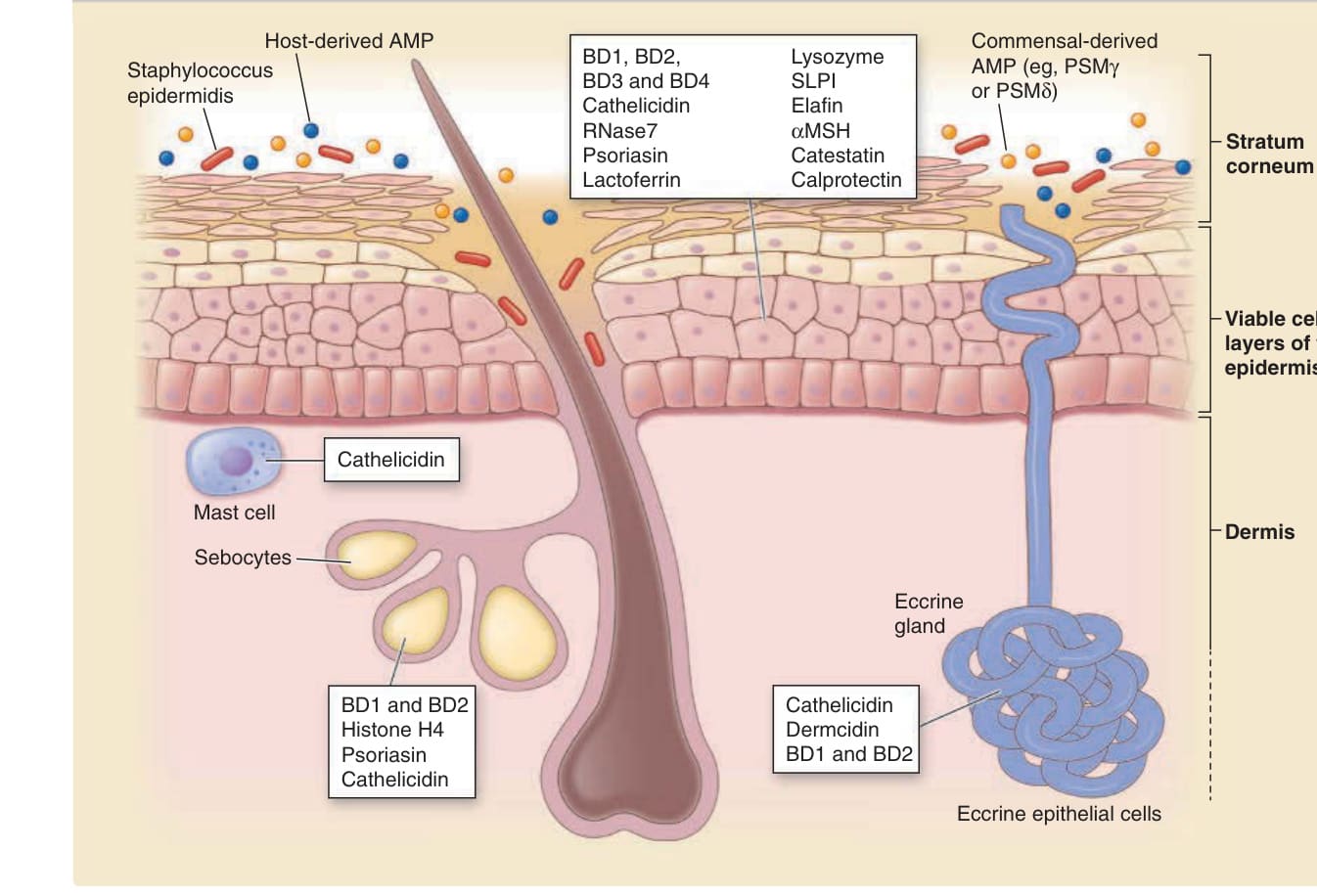
圖 14-12:由宿主細胞與共生細菌所產生的抗菌蛋白。AMP,抗菌蛋白 (antimicrobial proteins);BD,β-防禦素 (beta defensins);PSM,酚溶性調節素 (phenol soluble modulin);MSH,黑色素細胞刺激素 (melanocyte-stimulating hormone);SLPI,分泌型白血球蛋白酶抑制劑 (secretory leukocyte protease inhibitor)。(經 Springer Nature 許可改編自 Gallo RL, Hooper LV. Epithelial antimicrobial defence of the skin and intestine. Nat Rev Immunol. 2012;12:503.)
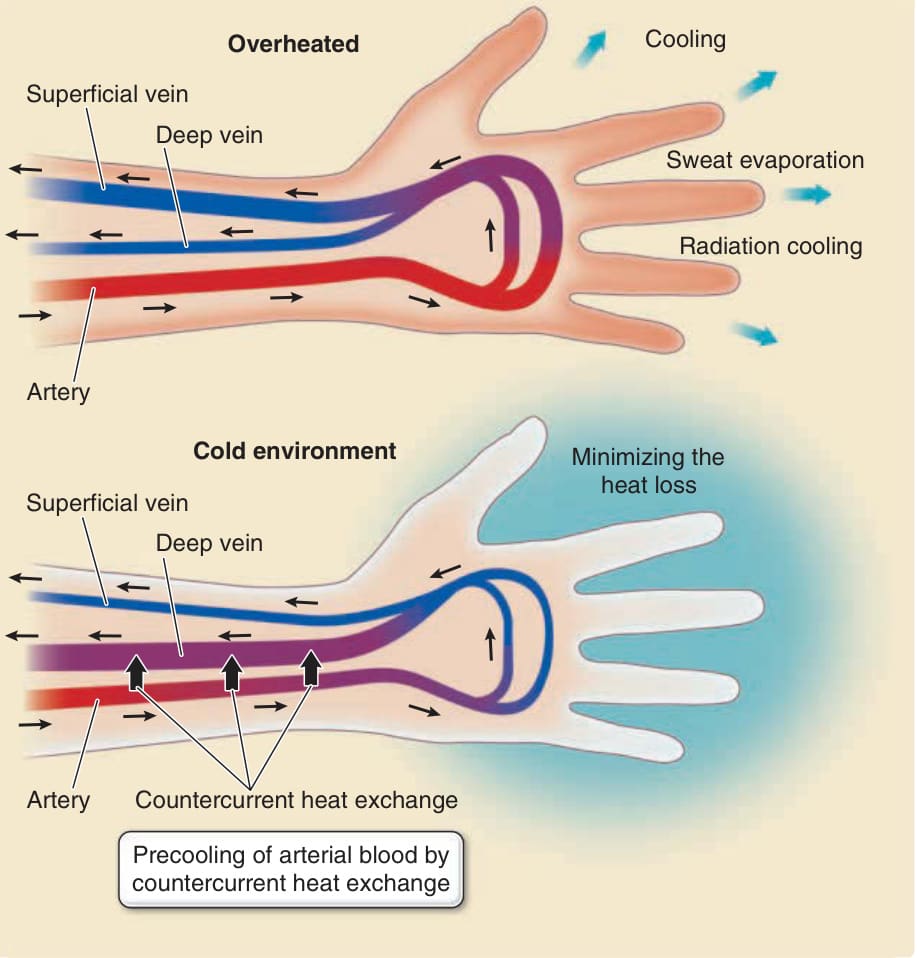
圖 14-13:皮膚中用於冷卻與維持熱量的血流控制。在炎熱環境中(上圖),淺表靜脈擴張,淺表靜脈中的血液主要經由體表外分泌汗液 (eccrine sweat) 的蒸發而冷卻。在寒冷環境中(下圖),淺表靜脈收縮,防止血液的熱量流失。傳入動脈與擴張的傳出深靜脈之間的逆流熱交換,在血液到達淺表血管之前預先冷卻動脈中的血液,這也防止了體表的熱量流失。

表 14-1:術語簡短註解 (Short Commentary on Jargons)
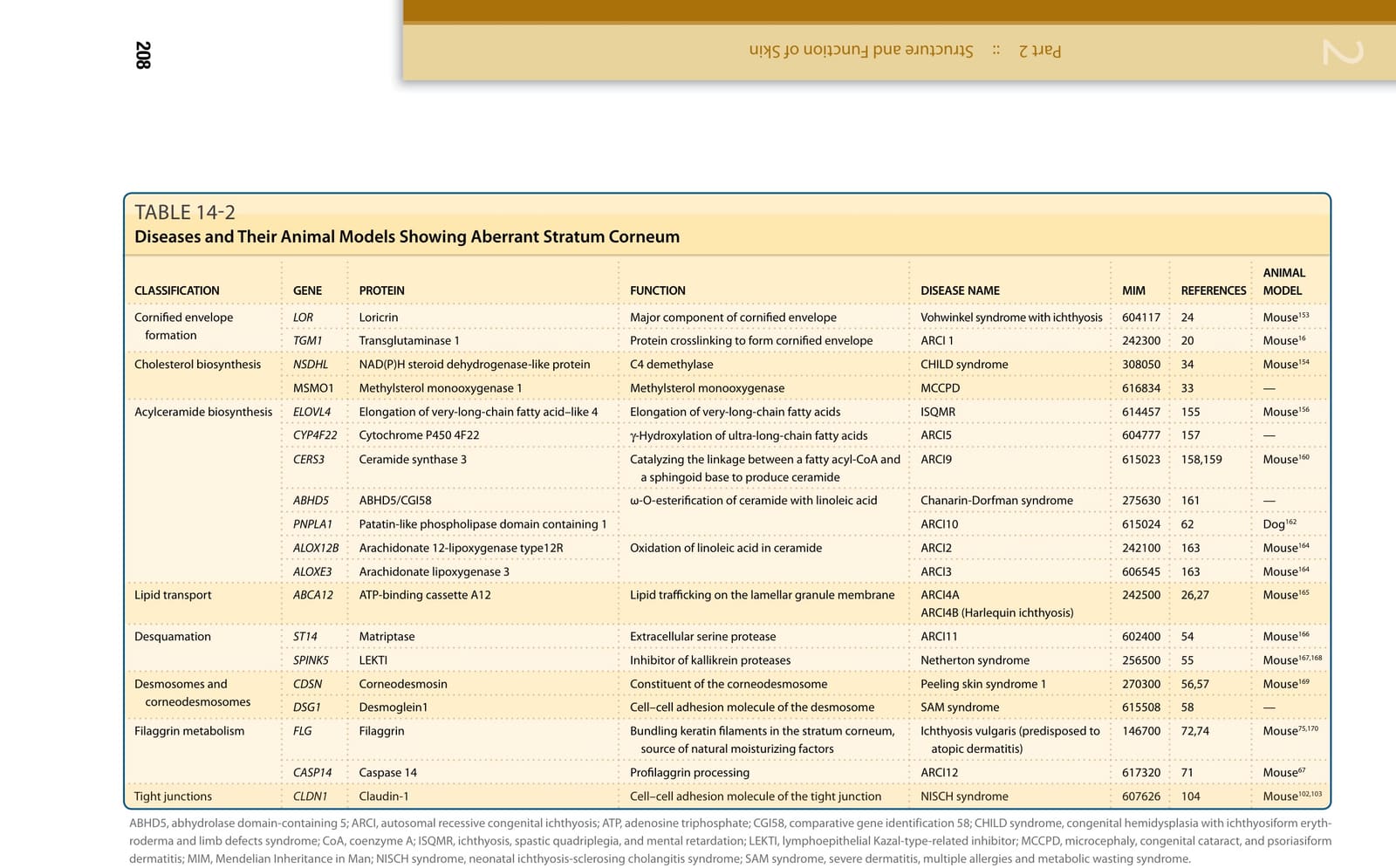
表 14-2:表皮屏障形成缺陷所造成的先天性疾病
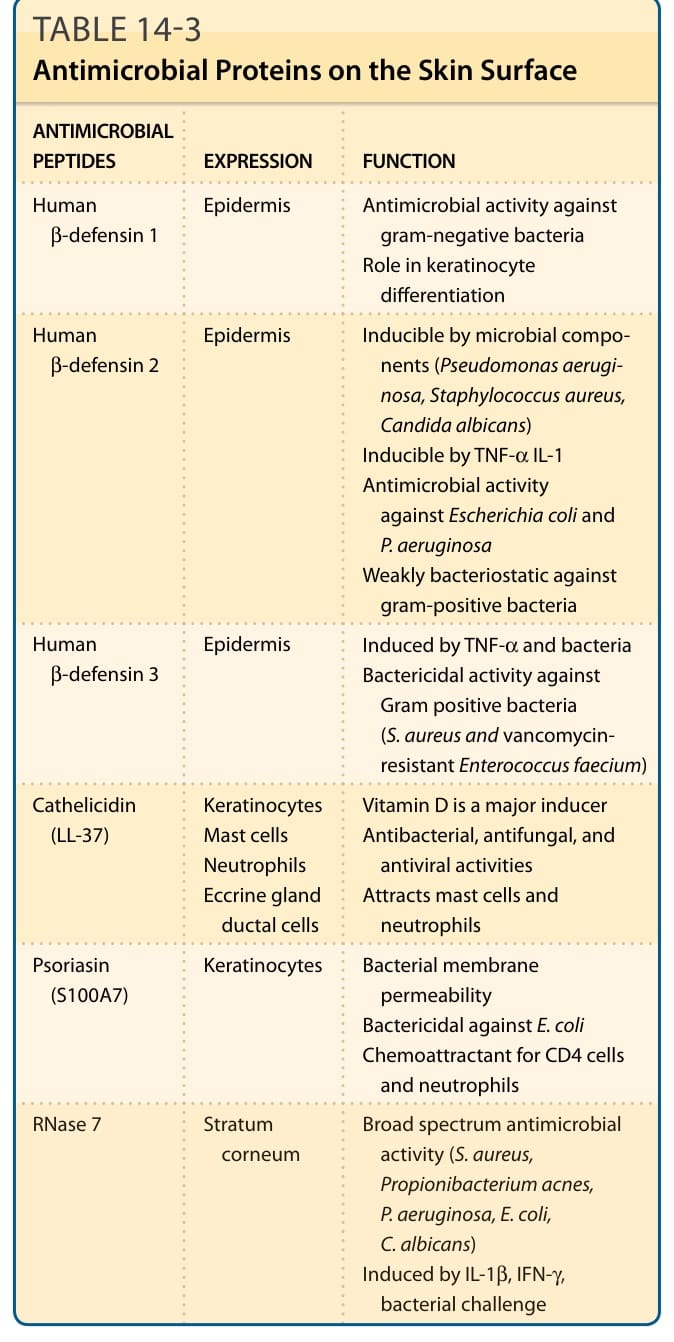
表 14-3:皮膚表面的抗菌蛋白 (Antimicrobial Proteins on the Skin Surface)
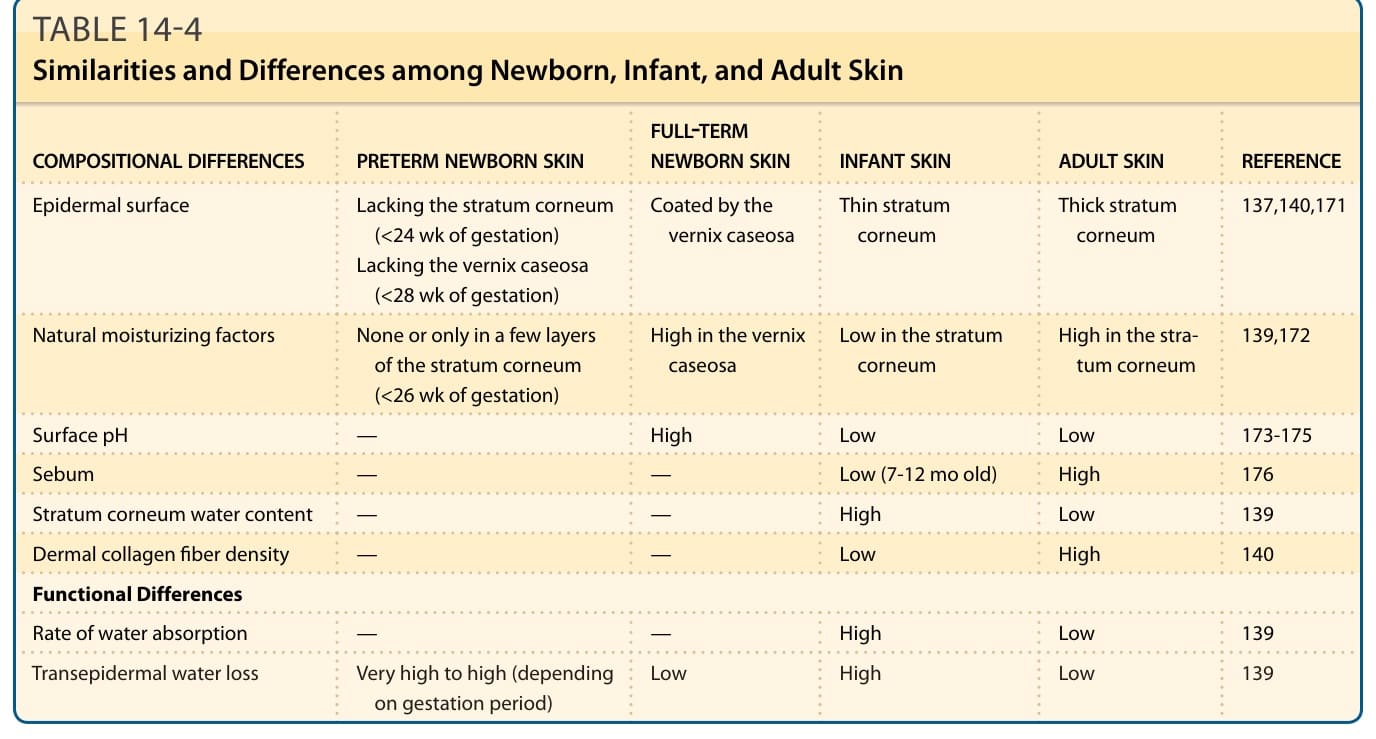
表 14-4:新生兒、嬰兒與成人皮膚之間的異同 (Similarities and Differences among Newborn, Infant, and Adult Skin)