Skin Barrier
2
AT-A-GLANCE
■ One of the most important functions of the skin is to form a barrier between the organism and the external environment.
■ The skin protects our bodies from physical damage caused by desiccation, physical stress, infection, overheating or heat loss, and ultraviolet (UV) irradiation.
■ The skin is covered by the epidermis, a cornified, stratified epithelial cellular sheet that is equipped with a barrier formed by the stratum corneum and tight junctions.
■ The stratum corneum is an air–liquid interface barrier on the body surface that prevents excessive water loss (inside–outside barrier) and the entry of harmful substances from the environment (outside–inside barrier).
■ The stratum corneum barrier is composed of corneocytes and intercorneocyte waterimpermeable lipid lamellae. Corneocytes are wrapped with cornified cell envelope and corneocyte lipid envelope and contain keratin filaments associated with filaggrin, which is degraded to natural moisturizing factors.
■ The tight junctions seal the intercellular space between neighboring cells at the second layer in the stratum granulosum and form a liquid–liquid interface barrier that limits molecular movement through the paracellular pathway.
■ Langerhans cells are located in the epidermis under the tight junction barrier in steady state but extend their dendrites to the outside tight junction barrier upon activation to capture external antigens at the tips of the dendrites.
■ Antimicrobial peptides, lipids, the acidic pH of the stratum corneum, and continuous daily desquamation (daily detachment of dead skin cells) control the skin microbiota and protect us from infection by bacteria, yeasts, fungi, and viruses
■ UV light is reflected from the stratum corneum and absorbed by urocanic acid and melanin molecules, which protect genomic DNA from UV irradiation damage.
■ Sweating, blood flow control, and heat storage in subcutaneous adipose tissue protect us from cold and overheating.
INTRODUCTION
The skin is the integument of vertebrates (Table 14-1). One of the key functions of the skin is to form a protective physical barrier between the body and the external environment. The limitation of molecular movement across the skin is largely dependent on the epidermis and especially the stratum corneum in mammals. The epidermis prevents the inward and outward passage of water, electrolytes, lipids, and proteins, as well as insults from chemicals, bacteria, fungi, virus, toxins, and allergens. Defects in the formation of the epidermal barrier cause various congenital diseases (Table 14-2). The skin protects the body from various external stresses. In protecting from heat and cold stresses, the skin maintains the internal organs at a certain constant temperature by regulating blood flow, sweat production, thermal storage in the fat layer, and thermogenesis in brown fat cells. The skin protects the body from physical stresses through coordination of the rigid surface armor of the stratum corneum; keratin cytoskeleton; and cell adhesions between keratinocytes, epidermal–dermal junctions, and dermal collagen fibers; these are discussed in Chaps. 15 and 60. Furthermore, the epidermis reflects and absorbs ultraviolet (UV) radiation from the sun to protect the genomic DNA of cells; this is particularly important in preventing carcinogenesis, which is discussed in Chaps. 19 and 20. Skin appendages, such as sweat and sebaceous glands, hairs, and nails, also have physical and chemical barrier functions; these are discussed in Chaps. 6, 7, and 8, respectively. The skin is also equipped with immunologic barriers to the innate and acquired immune systems. Antimicrobial proteins are a diverse group of proteins that form a chemical barrier against microorganisms on the surface of the epidermis. Toll-like receptors on keratinocytes detect pathogen-associated molecular patterns such as lipoprotein and peptidoglycans that are broadly shared by bacteria but distinguishable from host molecules and control the immune responses against the microorganisms. Dendritic cells within the skin govern the acquired immunologic and allergic responses to external insults. The mechanisms of acquired immunity that protect our skin are discussed in Chaps. 10 and 11.
Integument The tough outer protective layer of an animal or plant that demarcates the body from the external environment
Occlusive junction Cell–cell adhesive junction that seals the intercellular space and controls paracellular molecular movement. Invertebrates and vertebrates have different occlusive junction structures. The occlusive junction of invertebrates is called a septate junction, and that of vertebrates is called a tight junction.
Comparative Biology Comparative biology exploits natural variation and disparities among species or taxa to understand the patterns of life by investigating phylogenies, evolution, development, physiology, and genomics
Kelvin’s Tetrakaidecahedron Fourteen-sided polyhedron composed of eight hexagons and six squares, proposed in 1887 by William Thomson (Lord Kelvin) as the best shape for packing equal-sized cells together to fill the space using the minimal surface area of each cell142
Transepidermal Water Loss The loss of water that passes from inside to outside a body through the epidermis. Measurement of transepidermal water loss is useful to evaluate the stratum corneum barrier as the elevation rates of transepidermal water loss are basically in proportion to the level of barrier damage or formation.
Transepidermal Water Loss The loss of water that passes from inside to outside a body through
the epidermis. Measurement of transepidermal water loss is useful to evaluate the stratum corneum barrier as the elevation rates of transepidermal water loss are basically in proportion to the level of barrier damage or formation.
COMPARATIVE BIOLOGY OF THE EPIDERMAL BARRIER
The lipid bilayer cell membrane provides a fundamental barrier in unicellular organisms. The cell membrane compartmentalizes the cell from the external environment. Homeostasis of the cell is maintained within the compartment demarcated by the cell membrane in an energy-dependent manner. Because the lipid bilayer membrane is highly fragile, most unicellular organisms have an additional barrier structure outside of the cell membrane (ie, the cell wall) that functions as a type of armor (Fig. 14-1). The body’s barrier to multicellular organisms is analogous to its barrier to unicellular organisms. The cell membrane of unicellular organisms corresponds to the epidermis, which is the epithelial cellular sheet that delineates the surface of multicellular organisms (see Fig. 14-1). To form a barrier with a sheet of cells, prevention of the leakage of water and solutes through intercellular spaces is crucial. Thus, the intercellular spaces are sealed by occlusive junctions, such as septate junctions in arthropods and tight junctions in vertebrates (see Table 14-1). The epidermis is composed of a single layer of cells in cephalochordates and urochordates, but multilayered (stratified) cells in vertebrates (see Fig. 14-1). Because the epidermal cells are fragile when directly exposed to harsh external environmental factors, such as hypotonic fresh water, hypertonic
2
sea water, or dry air, the surface of the epidermis of higher organisms is covered by various external barriers. Examples of these barriers include the cuticles of arthropods, tunics of tunicates, mucous of fish and amphibians, and cornified cell layer (stratum corneum) of adult amphibians, reptiles, birds, and mammals (see Fig. 14-1).1
BASIC STRUCTURE OF THE EPIDERMIS
The human epidermis is a stratified epithelial cellular sheet, the uppermost part of which is cornified to form the stratum corneum. The viable (enucleated) cell layers of the epidermis consist of the stratum basale (basal cell layer), stratum spinosum (spinous cell layer), and stratum granulosum (granular cell layer) (Fig. 14-2). The stratum granulosum consists of at least three layers of flattened granular cells.2-5 From outside to inside, the layers are named SG1, SG2, and SG3, and tight junctions seal the intercellular spaces in the SG2 layer.3,5 The stratum corneum consists of dead cornified cells (corneocytes) and functions as an air–liquid interface barrier. The intercellular spaces of the stratum corneum are filled with water-impermeable lipid lamellae. Under the stratum corneum, cells are immersed in a water environment. The extracellular water environment of the epidermis is divided into two parts by the tight junction barrier, which is a liquid–liquid interface barrier (see Fig. 14-2). To form the stratum corneum, the SG1 cells terminally differentiate into corneocytes by filling their intercellular spaces with lipid lamellae. The multilayered structure of the epidermis is maintained, and the epidermal cells are continuously turned over. Epidermal cells proliferate only in the basal layer, differentiate with detachment from the basement membrane, move upward, become flattened at the stratum granulosum, form a tight junction at the SG2 layer, lose the tight junction at the SG1 layer, terminally differentiate into corneocytes, and detach from the top of the cornified layer as squamous scales. Continuous turnover of the epidermis enables foreign substances attached to the skin to be discarded with the daily detachment of scales.
STRATUM CORNEUM
The stratum corneum of human skin is the outermost barrier of the body’s surface. The stratum corneum in humans is approximately 10 to 20 µm thick and contains about 10 to 25 layers of cornified cells.6 The stratum corneum is directly exposed to air and protects the inner cells from damage by desiccation. It acts as a two-way barrier to prevent not only inward penetration of external molecules and microorganisms but also outward leakage of water and solutes.
207
2
2
Basic structure of the surface barrier of unicellular and multicellular organisms
Outer barrier
Occlusive junction
Connective tissue
Basement membrane
Lipid bilayer cell membrane
Multicellular organism Monocellular organism
A
Simple epithelia
KEY
Tight junction
Basement membrane
Mucus, tunic, etc.
Urochordate cephalochordate
Stratified epithelia
Stratum corneum
Mucus
Fish amphibian tadpole Amphibian adult, aves, reptiles, mammals
B
Vertebrate
Chordate
BASIC STRUCTURE OF THE STRATUM CORNEUM
BASIC STRUCTURE OF THE
STRATUM CORNEUM
The stratum corneum consists of stratified corneocytes and an intercorneocyte lipid-rich matrix (Fig. 14-3). The barrier function of the stratum corneum depends on both the protein-rich materials of the corneocytes and the intercellular lipid-rich matrix. Corneocytes are terminally differentiated dead keratinocytes that adhere to one another via proteinaceous cell–cell adhesion complexes called corneodesmosomes and the adhesive force of intercellular lipid lamellae (Fig. 14-4). On routine hematoxylin and eosin staining of paraffin-embedded skin sections, the stratum corneum shows a basket-weave structure, which
may cause the stratum corneum to be mistaken for a porous structure that does not have a barrier function; the basket-weave structure is, in fact, an artifact of the processing of specimens. During paraffin removal using xylene, intercorneocyte lipid is extracted, and the intercellular lipid-dependent adhesion between corneocytes becomes weakened. Because the corneodesmosomes make the lateral adhesions between corneocytes more stable than the apicobasal adhesions, the corneocyte layers detach from one another while maintaining their lateral adhesions, resulting in the formation of a basket-weave structure in paraffinembedded sections7,8 (Fig. 14-5). Proper fixation and staining procedures reveal the tightly packed, wellorganized structure of the stratum corneum in vertical sections of the skin2,9 (see Fig. 14-5).
209
2
Basic structure of the epidermis and major epidermal barriers
Physical assaults (dessication, irritation, UV irradiation, heat and cold shock)
Microbial assaults (bacteria, fungus, virus) Chemical assaults (irritants, allergens)
Stratum corneum
SG1
Stratum granulosum
SG2
SG3
Stratum spinosum
Stratum basale
Basement membrane
Heat control
KEY
Tight junction LEKTI KLKs
A
acidic
pH gradient
Stratum corneum
neutral
SG1
Stratum granulosum
SG2
SG3
Lamellar body
B
Scale desquamation
Cornification
Tight junction
Prevention of loss of water, solutes, and nutrients
Melanin cap
Cutaneous vasculature
Composition of the extracellular space
Desquamation
Intercellular lipid lamellae (Lipid-based air–liquid interface barrier)
Cell turnover
Cornification and intercellular lipid barrier formation
Extracellular aqueous environment outside tight junction
TJ barrier (Liquid–liquid interface barrier)
Extracellular aqueous environment inside tight junction
210
Basic structure of the stratum corneum
Corneocyte Paracellular route of permeation Intercellular lipid lamellae
Protein-based cornified cell envelope
Ceramide coat of cornified envelope (corneocyte lipid envelope)
Lipid-enriched extracellular matrix with lipid lamellae
FORMATION OF A CORNIFIED CELL ENVELOPE AND CORNEOCYTE LIPID ENVELOPE
FORMATION OF A
CORNIFIED CELL ENVELOPE
AND CORNEOCYTE LIPID
ENVELOPE
Granular layer cells terminally differentiate into corneocytes to form the stratum corneum. During cornification, a cornified cell envelope consisting of a 10-nm-thick layer of highly cross-linked insoluble proteins is formed beneath the plasma membrane,10 and the lipid bilayer of plasma membrane is replaced by a 5-nm-thick layer of acylceramides, which is called corneocyte lipid envelope11-13 (Fig. 14-6). The formation of a cornified cell envelope and corneocyte lipid envelope is described as follows14,15 (see Fig. 14-6). Envoplakin, periplakin, and involucrin expressed in granular layer cells associate with the inner surface of the plasma membrane in a calciumdependent manner and are cross-linked to one another by transglutaminase 1.16 Transglutaminase 1 also cross-links to other membrane-associated proteins and desmosomal proteins, fixing the cell junctions and associated cytoskeletons to the proteinaceous scaffold. The involucrin-envoplakin-periplakin–based scaffold eventually forms a monomolecular layer along
2
the entire inner surface of the plasma membrane.10,17-19
Defects in transglutaminase 1 are known to cause congenital ichthyosis (see Table 14-2).20
Before the scaffold covers the inner surface of the plasma membrane, lamellar bodies are produced from the Golgi complex and become fused to the plasma membrane. The limiting membrane of the lamellar bodies is rich in acylceramides composed of omega-hydroxylated ultra-long-chain fatty acids. The fusion of the limiting membrane with the plasma membrane gradually increases the amount of acylceramides within the lipid bilayer of the plasma membrane. The acylceramides become covalently bound to the outer surface of the scaffold of the cornified envelope by the membrane-anchored form of transglutaminase 1.21,22 The acylceramides eventually replace the plasma membrane, which is called the corneocyte lipid envelope11,12 (see Fig. 14-6). The involucrin-envoplakin-periplakin–based scaffold is further reinforced in the later stage of cornification. On the inner surface of the scaffold, loricrin molecules are translocated from the cytosol and covalently cross-linked onto the scaffold to build the cornified cell envelope. Loricrin, an insoluble protein that may contribute to the water resistance of the cornified cell envelope, eventually becomes the major component of the cornified cell envelope. Varying amounts of small proline-rich proteins with minor amounts of other proteins (eg, repetin, trichohyalin, cystatin A, elafin, and late envelope proteins) are also cross-linked to the inner surface of the cornified envelope.23 Defects in loricrin are known to cause congenital ichthyosis (see Table 14-2).24
FORMATION OF THE LAMELLAE OF INTERCORNEOCYTE LIPIDS
FORMATION OF
THE LAMELLAE OF
INTERCORNEOCYTE LIPIDS
Keratinocytes of the stratum granulosum develop a specific system of lamellar bodies that allow secretion of intercorneocyte lipid lamellae.25 Lamellar bodies are produced from the Golgi complex and stored within the cytoplasm in SG3 cells as intracellular vesicles. Lamellar bodies are enriched in polar lipids, glycosphingolipids, free sterols, and phospholipids. ABCA12 functions in cellular lipid trafficking on the limiting membrane of lamellar bodies, in which severe defects cause Harlequin ichthyosis (see Table 14-2).26,27 The lamellar bodies may also contain proteins, such as hydrolytic enzymes to modify lipids, corneodesmosins to modify corneodesmosomes, antimicrobial peptides, and proteases and protease inhibitors to control desquamation. The contents of the lamellar bodies are delivered into the extracellular milieu through exocytosis occurring via the apical cell membrane, most probably in the SG2 cells and SG1 cells, to fill the extracellular space surrounding the SG1 cells28 (see Fig. 14-2).
211
2
E
De
A B
LB
200 nm
C D
200 nm
E
CD
200 nm
Lipid lamellae
The lipids exocytosed from the lamellar bodies are subsequently organized into a characteristic lamellar structure that lies parallel to the cornified cell envelope during cornification of the SG1 cells (see Fig. 14-4). The covalently bound acylceramides of the corneocyte lipid envelope act as a scaffold allowing regular lamellar formation of the intercellular lipids (see Figs. 14-4 and 14-6).22 After extrusion of the lamellar bodies into the extracellular spaces, the polar lipids are enzymatically converted into nonpolar products. Glycosphingolipids are hydrolyzed to generate ceramides, and phospholipids are converted into free fatty acids. These lipids form the intercellular lamellar component of the stratified lipid bilayer, a very dense structure packed into the interstices of corneocytes, thus forming a waterimpermeable barrier.
212
LIPID COMPOSITION OF THE STRATUM CORNEUM
LIPID COMPOSITION OF
THE STRATUM CORNEUM
Intercellular lipids are indispensable in the formation of the permeability barrier of the stratum corneum. The major classes of lipids in the stratum corneum are cholesterol, free fatty acids, and ceramides.29
Cholesterol forms part of the plasma membrane in viable cell layers in the epidermis and part of the intercellular lipid lamellae in the stratum corneum. Whereas basal-layer keratinocytes are capable of resorbing cholesterol from the circulation, epidermal keratinocytes actively biosynthesize cholesterol and free fatty acids (Fig. 14-7). The majority of the cholesterol in the epidermis is synthesized
A
B
C
in situ from acetyl coenzyme A (acetyl CoA).30 The rate-limiting step in cholesterol biosynthesis is catalyzed by hydroxymethylglutaryl CoA (HMG CoA) reductase (see Fig. 14-7). Epidermal cholesterol biosynthesis is upregulated during barrier repair.31 The cholesterol biosynthesis pathway in the epidermis is also important for the production of vitamin D.32
2
The metabolic intermediate of cholesterol biosynthesis, 7-dehydrocholesterol, is converted to previtamin D in a photolytic reaction by UVB radiation; this is followed by thermal isomerization to form vitamin D3. Deficiency in the enzymes of NSDHL and MSMO1 corresponding to cholesterol biosynthesis causes inflammatory ichthyotic skin lesions through the accumulation of toxic intermediates in CHILD (congenital hemidysplasia with ichthyosiform erythroderma and limb defects) syndrome and MSMO1 deficiency, respectively33-35 (see Table 14-2). The skin contains free fatty acids, as well as fatty acids bound in triglycerides, phospholipids, glycosylceramides, and ceramides. Although biosynthesis of cholesterol originates from HMG-CoA, free fatty acid synthesis originates from malonyl-CoA produced from acetyl-CoA via acetyl-CoA carboxylase (see Fig. 14-7). Saturated and monounsaturated fatty acids are synthesized in the epidermis. However, not all fatty acids can be synthesized by the epidermis or the human body. Fatty acids unable to be produced by the human body are called essential fatty acids. Essential fatty acid deficiency (EFAD), caused by unusual diets or malabsorption in humans or experimentally induced in rodents, leads to rough, scaly, and erythematous skin with severe permeability barrier defects of the epidermis.36,37
Ceramides consist of long-chain amino alcohols, called sphingoid bases, linked to a fatty acid via an amide bond (see Fig. 14-7). Ceramides are synthesized by serine palmitoyltransferase and hydrolysis of both glucosylceramide and sphingomyelin in the epidermis. Whereas ceramides are a minor lipid component in the mammalian body (<10% of cholesterol or phospholipids), they are a major lipid component in the stratum corneum, accounting for 30% to 40% of lipids by weight.29 Such a high ceramide content is only observed in the stratum corneum and not in the stratum granulosum, stratum spinosum, or stratum basale. This distribution indicates that ceramide biosynthesis is spatiotemporally controlled and highly activated in the uppermost keratinocytes under terminal differentiation to corneocytes. Among the epidermal ceramides with marked molecular heterogeneity, acylceramide is essential not only for the formation of the corneocyte lipid envelope, as described earlier, but also for the proper organization of intercorneocyte lipid lamellae and thereby the barrier function of the stratum corneum.38-42 Acylceramide is synthesized via esterification of omegahydroxyceramide with linoleic acid. Acylceramide is an unusual ceramide, the N-acyl chain of which is composed of omega-hydroxylated ultra-long-chain fatty acids (see Fig. 14-7). It has been suggested that the ultra-long-chain fatty acid of acylceramide spans a bilayer of lipid lamellae to link the two membranes together in the lipid lamellae and thus serves as a molecular rivet to form stratified lipid lamellae.43 Deficiencies of the biosynthesis pathway of acylceramide cause several types of congenital ichthyosis44 (see Table 14-2).
213
2
Formation of the cornified cell envelope, corneocyte lipid envelope, and intercorneocyte
lipid lamellae
Differentiation from stratum granulosum cells to corneocytes
5 3 4
Conversion from peripheral cell membrane to corneocyte lipid envelope
Conversion from desmosome to corneodesmosome Lipid lamellar formation
Desmosome Corneodesmosome
Intercellular space surrounding SG1 cell
Lipid bilayer cell membrane
Acylceramide
Intercorneocyte lipid lamellae Corneocyte lipid envelope (~5 nm thickness)
TG1
Cornified cell envelope (~10 nm thickness)
Loricrin and Small prorine-rich proteins Secretion of the lamellar body contents
2
Lipids, proteases, protease inhibitors, and antimicrobial molecules Lamellar body
Keratin filaments
1
Formation of lamellar body from Golgi
Limiting membrane rich in acylceramide
Involucrin Envoplakin Periplakvin
Lattice structure of keratin filaments linked with filaggrin
6
Formation of cornified envelope by cross-linking scaffold proteins via transglutaminzese 1
Loricrin granule
CORNEODESMOSOMES
CORNEODESMOSOMES
Desmosomes are the major cell–cell adhesion structure in the granular, spinous, and basal cell layers. Corneodesmosin is secreted by lamellar bodies into the extracellular spaces surrounding the SG1 cells (see Figs. 14-2 and 14-6) and then integrated into the desmosomes, resulting in the formation of corneodesmosomes (ie, specific cell adhesion structures between corneocytes)45 (see Fig. 14-4). Adhesion between corneocytes is dependent on both corneodesmosomes and intercellular lipid lamellae. Corneodesmosomes are degraded in the outer layers
214
of the stratum corneum, and the outermost corneocytes are detached, one by one, from the top layer of the stratum corneum; this process is called desquamation.46
The major proteases involved in the degradation of corneodesmosomes are serine proteases belonging to the kallikrein group. Kallikreins 5, 7, and 14 are known to exist in the stratum corneum. These kallikreins are also secreted by lamellar bodies into the extracellular spaces surrounding the SG1 cells (see Fig. 14-2). Kallikreins are produced as inactive precursors and activated via proteolytic conversion by kallikreins themselves (autoactivation) or by matriptase, a transmembrane serine protease.47,48 The protease activity
2
Synthetic pathways and key enzymes for cholesterol, free fatty acids, ceramides, and acylceramides
Acetyl-CoA
HMG-CoA synthase
HMG-CoA reductase HMG-CoA
Mevalonic acid
Squalene
Lanosterol
Acetyl-CoA carboxylase
Cholesterol synthesis
Deficiencies in enzymes
Dihydrolanosterol
CHILD syndrome MCCPD
NSDHL / MSMO1 NSDHL / MSMO1
Zymosterol
Dehydrodesmosterol
Zymostenol
7-Dehydrocholesterol
7-Dehydrocholesterol reductase Ultraviolet B
Desmosterol Previtamin D
Cholesterol
Fatty acid synthase
Ceramide synthesis
ISQMR ARCI5
Fatty acids ULC-fatty acids
Malonyl-CoA
Phospholipase A2
Phospholipids
Serine + Palmitoyl-CoA
Ceramide synthase
Sphinganine
Food intake (Essential fatty acids)
Linoleic acid
Ceramides
ARCI9
Chanarin-Dorfman syndrome ARCI10 ARCI2 ARCI3
Acylceramides
of the activated kallikreins is thought to be inhibited by the direct binding of lymphoepithelial Kazal-typerelated inhibitor (LEKTI), which is also secreted into the intercellular spaces via lamellar bodies.49 The production of kallikreins as inactive precursors, and the inhibition of their protease activity by LEKTI, are considered to prohibit the degradation of corneodesmosomes in the lower layer of the stratum corneum and thus prevent premature desquamation.50 LEKTI binding to kallikreins has been shown to be pH dependent. The lower pH of the intercellular spaces in the outer layers of the stratum corneum is thought to facilitate the dissociation of kallikreins from LEKTI and the kallikrein-dependent degradation of corneodesmosomes (see Fig. 14-2).51-53 The outermost corneocytes eventually detach from the skin.
Congenital defects of matriptase induce congenital ichthyosis showing hyperkeratosis and impaired degradation of corneodesmosomes, probably caused by insufficient activation of kallikrein proteases.54 In contrast, congenital defects of LEKTI induce Netherton syndrome, in which entire layers of the stratum corneum are prone to being peeled off, probably caused by the enhanced degradation of corneodesmosomes by kallikreins.55 Such detachment of whole layers of the stratum corneum is also observed in the congenital defects of corneodesmosin or desmoglein 156-58 (see Table 14-2). Patients prone to total detachment of the stratum corneum are predisposed to various allergic conditions, probably via increased allergen penetration through the defective skin barrier and facilitation of percutaneous sensitization.5,59
215
2
KERATIN CYTOSKELETON AND FILAGGRIN
KERATIN CYTOSKELETON
AND FILAGGRIN
Filaggrin is the major protein of the stratum corneum. Various mutations in the filaggrin gene predispose patients to atopic eczema, allergic rhinitis, food allergies, and asthma complicated with eczema.60-63
In humans, filaggrin is expressed only in cornified stratified epithelia; therefore, it is expressed in skin but not in the esophagus or airways.64 Profilaggrin is expressed in the stratum granulosum and forms intracellular aggregates known as keratohyalin granules. Under cornification, profilaggrin is processed by various proteases into mature filaggrin monomers63
(Fig. 14-8). Filaggrin monomers bundle keratin
Multi-domain structure of the stratum corneum and multi-step maturation/degradation of filaggrin
H2O Hg2+, Cr2+, K+
Fluorescein Allergens
Corneocytes of the upper SC
Caspase 14 plus additional peptidases
Bieomycin hydrolase
Corneocytes of the middle SC
Intermediate species
PAD1 and PAD3 TGMs
Corneocytes of the lower SC
Formation of keratin-based matrix
Profilaggrin
Ca2+
Dephosphorylation
Proteolysis Multiple protease
Cornification
UV
Water-soluble small molecules permeate into the upper layers of the stratum corneum where protein structure is proteolytically degraded.
Amino acids
In the middle layers of the stratum corneum, filaggrin degraded into natural moisturizing factors.
+UCA +PCA
NMF
In the lower layers of the stratum corneum, filaggrin monomers bundle keratin filaments.
Nucleus
Nuclei and intracellular organella are degraded.
Profilaggrin proteolytically degraded into mature filaggrin monomers.
A and B domain of profilaggrin move into the nucleus.
Mature filaggrin monomers Nucleus
Keratohyalin granules
Keratin IFs
Stratum granulosum
Nucleus
Profilaggrin forms keratohyalin granules.
KEY
Profilaggrin
A domain Filaggrin
B domain Free amino acid C-terminal domain
216
intermediate filaments and may form a lattice structure in the lower layer of the stratum corneum.65,66
The lattice structure can be observed under electron microscopy. In the later stage of cornification, keratin filaments bundled with filaggrin become cross-linked to the cornified cell envelope. The dead cornified cells lose most of their intracellular organelles and consist mostly of bundled intermediate filaments covalently attached to, and enclosed within, the cell envelope.
FILAGGRIN DEGRADATION AND NATURAL MOISTURIZING FACTORS
FILAGGRIN DEGRADATION
AND NATURAL
MOISTURIZING FACTORS
Filaggrin monomers bundle keratin filaments in the innermost layers of the stratum corneum. Because new corneocytes are continuously supplied at the bottom and old corneocytes are desquamated from the surface of the stratum corneum, each corneocyte gradually moves to the upper layers. During this upward movement of the corneocyte, intracellular filaggrin molecules are degraded into free amino acids and their derivatives by proteases, including caspase 14 and bleomycin hydrolase. Representative examples of the derivatives are trans-urocanic acid catalyzed from histidine by histidase and pyrrolidone carboxylic acid catalyzed from glutamine.63,67-69
The natural moisturizing factors of the stratum corneum are composed primarily of these amino acids and their derivatives, together with lactic acid, urea, citrate, and sugars.61,70 Deficiency of caspase 14 causes a defect in the proper degradation of filaggrin and the resultant phenotype of ichthyosis71
(see Table 14-2). Haploinsufficiency of filaggrin, because of various heterozygous mutations in the gene encoding filaggrin, causes ichthyosis vulgaris, a dry skin condition,72
and predisposes patients to atopic dermatitis and various allergic diseases accompanied by eczema (eg, food allergy and asthma)73,74 (see Table 14-2). The decrease in filaggrin induces a decrease in natural moisturizing factors, a disturbance of the lattice structure of the keratin filaments in the innermost layers of the stratum corneum, and a probable increase in the incidence of barrier breakage of the stratum corneum.75 Such a weakness in the stratum corneum barrier may increase the chance of external allergens penetrating into the skin and the risk of percutaneous sensitization against various allergens. Most patients with severe stratum barrier defects, such as those with Netherton syndrome, have multiple allergies. In contrast, most patients who have mutations in the filaggrin gene basically develop ichthyosis vulgaris, and only some of these patients develop allergic diseases. This suggests that the barrier deficiency induced by filaggrin mutations is moderate and that the development of allergic diseases in these patients is modulated by various additional factors, such as genetic differences in immune reactions,76 differences in the
2
skin microbiota,77-79 and environmental factors such as low air humidity80 and air pollution.81
MULTIDOMAIN STRUCTURE OF THE STRATUM CORNEUM
MULTIDOMAIN STRUCTURE
OF THE STRATUM
CORNEUM
Conventional cell biological analyses of the stratum corneum have been hampered by the insolubility of covalently cross-linked proteins. To overcome this difficulty, the stratum corneum has been analyzed by various special methods, such as cryoelectron microscopy, immunoelectron microscopy, Raman spectromicroscopy analysis, and time-of-flight secondary ion mass spectroscopy.82-85 Based on the results of these analyses, the stratum corneum is considered to be divisible into several different parts with different characteristics, most likely into three parts (the upper, middle, and lower layers). For example, water-soluble small molecules soak in and out at the upper part of the stratum corneum, where the protein-based structure of the corneocytes is proteolytically degraded, and the corneocytes are thought to function as a sponge.83,85 Further permeation of water-soluble small molecules into the middle part of the stratum corneum is rather limited, indicating that permeation barriers exist within the middle part of this structure.83 The middle part of the stratum corneum also has a large potential to absorb and hold water, likely corresponding to the enrichment of natural moisturizing factors, which are mostly produced by the degradation of mature filaggrin.63,82 The corneocytes of the lower part of the stratum corneum show an intracellular lattice structure of keratin filaments, integrated by mature filaggrin monomers, which is believed to contribute to the physical strength of the stratum corneum84 (Fig. 14-9). The stratum corneum is a pile of “dead” corneocytes that lack any energydependent activities, such as adenosine triphosphate– dependent active transport, translation of proteins, and mRNA transcription. It is still largely unknown how the stratum corneum maintains homeostasis using the distinct features in its upper, middle, and lower layers.
PERCUTANEOUS ABSORPTION THROUGH THE STRATUM CORNEUM
PERCUTANEOUS
ABSORPTION THROUGH
THE STRATUM CORNEUM
Knowledge of how external molecules penetrate the epidermis is important not only for a better understanding of the pathogenesis of various skin diseases but also for the development of drug delivery for topical treatments. The paracellular route is a major pathway by which external molecules can penetrate the stratum corneum (see Fig. 14-3). Other routes of penetration into the skin are by the sebaceous glands or hair follicles, but the exact mechanisms of these pathways remain to be elucidated.86
217
2
Tight junction barrier and cell polarity in simple and stratified epithelial cellular sheets
Outside TJ
Outside TJ
Simple epithelium Cornified epidermis
Apical membrane
TJ
Basolateral membrane
Paracellular pathway Transcellular pathway
Inside TJ
Cell membrane
TJ strands
100 nm
Intercellular space
Stratum granulosum Stratum corneum
SG1
SG1 SG1
SG2
SG2 SG2
SG3
SG3 SG3
Inside TJ
Claudins
N C
Occludin
N
C
Occlusive patches used in patch testing affect the intercorneocyte lipid lamellae and increase the permeation of solutes and solvents through the stratum corneum.87 After passing through the stratum corneum, small molecules such as haptens can easily permeate the dermis, probably through the transcellular pathway (see Fig. 14-9). In contrast, large peptide antigens, such as the antigens of house dust mites or egg albumin, not only find it difficult to pass
218
through the stratum corneum barrier but are also prohibited from further permeation into the skin by the tight junction barrier. Recent studies revealed that a defective stratum corneum barrier in early childhood is associated with the development of various allergic diseases, such as atopic dermatitis, allergic asthma, and food allergies, probably via enhanced antigen penetration through the stratum corneum and percutaneous sensitization.74,88-91
TIGHT JUNCTIONS
Multicellular organisms use epithelial cellular sheets to separate their body from the external environment or their organs from one another. Epithelia serve as selective permeability barriers to fluids that differ in chemical composition (Fig. 14-10). The occluding junctions are specialized intercellular adhesion complexes that are crucial for limiting molecular movement through the paracellular pathway (see Fig. 14-10) because they seal the intercellular spaces of the epithelial cellular sheets.92 Tight junctions facilitate this barrier in vertebrates. In the gut of humans, for example, gut epithelia equipped with tight junction barriers separate body fluids from the fluid of the gut lumen.
STRUCTURE OF TIGHT JUNCTIONS
STRUCTURE OF TIGHT
JUNCTIONS
In simple epithelia, the tight junctions are located at the apical-most part of the apical junctional complex93 (see Fig. 14-9). The apical junctional complex consists of tight junctions, beltlike adherens junctions, and desmosomes. The cell membrane is divided into two parts at the tight junctions—the apical cell membrane and basolateral cell membrane—both of which has a different composition of membranous proteins. Tight junctions
2
consist of tight junction strands, which are mainly composed of four-transmembrane proteins of claudins on the plasma membrane that probably function as a zip lock to seal the adjacent plasma membrane, thereby forming a barrier against molecular movement through the paracellular pathway (see Fig. 14-9).94,95 Other major components of tight junctions are the transmembrane proteins of occludin, junctional adhesion molecule A (JAM-A), tricellulin, and angulins, and the intracellular scaffold proteins of ZO-1, 2, and 3.96,97 Tight junctions do not simply form an impermeable barrier but also serve as ion- and size-selective barriers that vary in tightness depending on the cell type and the composition of their structural adhesive molecules.97-99
FUNCTION OF TIGHT JUNCTIONS
FUNCTION OF TIGHT
JUNCTIONS
Observations of normal human trunk skin and mouse ear, abdominal, and back skin revealed that only the single-layer cells of the SG2 layer are equipped with tight junctions and show apical and basolateral cell membrane polarity in the epidermis (see Figs. 14-2 and 14-9).100,101 The extracellular spaces of the epidermis are divided into two parts by the tight junction barrier.5 The SG2 and SG1 cells secret various molecules (ie, lipids and proteases) via lamellar body secretions from
Spatial location of the stratum corneum, tight junctions, and Langerhans cells
TJ barrier
Ag
Ag uptake Stratum corneum
SG1
Stratum granulosum
SG2
SG3
Stratum spinosum
Outside of TJ barrier
Inside of TJ barrier
Steadystate LC Actiated LC
Stratum basale
Basement membrane
219
2
their apical cell membrane into the extracellular spaces surrounding the SG1 cells, which are outside the tight junction barrier and are probably segregated from the environment inside the tight junction barrier (see Fig. 14-2). A lack of claudin-1, a major claudin of the epidermis, causes leakage of the epidermal tight junction barrier and deficient formation of the stratum corneum barrier in mice, resulting in early neonatal death (probably caused by water loss from the body surface).102
Decreased claudin-1 expression in mice induces an ichthyosis phenotype and skin inflammation.103 A lack of claudin-1 in humans results in a very rare congenital disease called neonatal ichthyosis sclerosing cholangitis syndrome104 (see Table 14-2). These observations indicate that a functional tight junction barrier is crucial for proper formation of the stratum corneum barrier and maintenance of epidermal homeostasis.
INTERACTION BETWEEN TIGHT JUNCTION BARRIERS AND INTRAEPIDERMAL IMMUNE-MEDIATED CELLS
INTERACTION BETWEEN
TIGHT JUNCTION BARRIERS
AND INTRAEPIDERMAL
IMMUNE-MEDIATED CELLS
Langerhans cells are antigen-presenting cells that are found in the epidermis and have multiple dendrites. They stay below the tight junction and project their dendrites upward in their steady state in both human and mice epidermis.101,105 After activation, Langerhans cells extend their dendrites beyond the tight junction barrier and capture external antigens that penetrate through the stratum corneum (see Fig. 14-10). During this process, new tight junctions are formed between Langerhans cell dendrites and keratinocytes of the SG2 layer, enabling the uptake of external antigens without disturbing the barrier function of the tight junctions. The Langerhans cells migrate out from the epidermis, present the captured antigens to T cells in draining lymph nodes, and are considered to induce humoral immunity to the captured antigens.101,106
γ-δ T cells are a minor subpopulation of T cells that express T-cell receptor γ and δ. In the human epidermis, γ-δ T cells are rare; however, a large proportion of T cells are γ-δ T cells in mice epidermis.107-110 The dendrites of γ-δ T cells have been shown to dock with the basal side of the tight junction-bearing cells in mice epidermis, forming immunologic synapses that polarize and anchor T cell projections at keratinocytes.111
SHAPE OF KERATINOCYTES FOR SKIN BARRIER HOMEOSTASIS
SHAPE OF KERATINOCYTES
FOR SKIN BARRIER
HOMEOSTASIS
Corneocytes form according to a regular stacking structure, with regular zigzag interdigitation patterns
220
between adjacent cell columns (see Fig. 14-5B). This regularly interdigitated stacking structure is most apparent in the skin of rodent ears but is also observed in the skin of other body regions in rodents, as well as in human skin.112 Classic scanning electron microscopic studies of the skin surface corneocytes of the murine ear and a recent immunofluorescent microscopic study of the tight junction-bearing granular layer cells of the murine ear revealed that the basic shape of the cell is a flattened variation of Kelvin’s tetrakaidecahedron (see Table 14-1) in the stratum corneum and stratum granulosum4,113,114 (Fig. 14-11). The regular interdigitation pattern of the corneocytes is believed result from the regular interdigitated stacking of the flattened tetrakaidecahedron cells (see Figs. 14-5B and 14-11C). Computational simulation and in vivo imaging studies suggested that this regular structure is formed at the granular layer by transformation of the three-dimensional shape of cells supplied from the spinous layer, although the precise molecular mechanisms underlying the cell shape change are still unknown.4,115-117 Only a single layer of cells (SG2 layer cells) form tight junctions at the apical edges of lateral cell–cell contacts. Therefore, tight junctions are formed at the edges of tetrakaidecahedron cells (see Fig. 14-11). When a tight junction-forming cell (an old cell) is replaced by a new cell located just beneath the old cell, the new cell first forms a new tight junction polygon beneath the old cell. Thus, each tight junction polygon becomes temporally double-edged during cell replacement (see Fig. 14-11). After that, the tight junction polygon of the old cells disappears, and eventually, the old cell is extruded to the outside of the tight junction barrier (the outside of the SG2 cell layer that forms the tight junction barrier). The temporal formation of the double-edged tight junction polygon at each cell turnover site maintains the homeostasis of the tight junction barrier.4 The regular interdigitated stacking of the flattened tetrakaidecahedron cells enables not only the temporal formation of the double-edged tight junction polygon but also to maintain the spatial relationship between cells and desmosomal cell–cell adhesions during cell turnover that may be responsible the physical strength of the epidermis.4 The molecular mechanisms that restrict tight junction-forming activity to the SG2 cell layer in the epidermis, the spatiotemporal regulation of cell turnover while maintaining tight junction barrier homeostasis, and the manner in which the tetrakaidecahedron cell shape is produced are currently unknown.
ANTIMICROBIAL BARRIER
ANTIMICROBIAL PROPERTY OF THE SKIN SURFACE
ANTIMICROBIAL PROPERTY
OF THE SKIN SURFACE
The skin constantly encounters microbial pathogens. Control of the skin microbiota is important to prevent bacterial and fungal infections. Continuous
2
Three-dimensional cell shape and a model for cell replacement in the epidermis
Tight junction–bearing cells in the stratum granulosum
Corneocytes at the surface of the stratum corneum
Schema of the epidermis based on the tetrakaidekahedron cell shape
Stratum granulosum
SG1
SG1
TJ
SG2
TJ
SG2
SG3
SG3
A
Kelvin’s tetrakaidecahedron
Flattened Kelvin’s tetrakaidecahedron
Stack of flattened Kelvin’s tetrakaidecahedron
SG2 SG2 SG2
SG2
B
SG2
SG2
SG2
SG2
SG2
2 3
2 3
Stratum corneum
7
7
1
1
4
4
6
6
SG1
S
5
5
SG2
TJ
TJ
S
SG3
S
Double-edged TJ polygon in the epidermis
KEY
Tight junction
C
SG2
SG2
SG2
SG2
SG2
SG2
Time-dependent TJ replacement and cell extrusion to the outside TJ barrier
D
221
2
turnover of epidermal cells by desquamation prohibits colonization of microorganisms on the skin. The physical properties of the skin surface itself prevents bacterial growth, that is, a low carbohydrate and water content and a weakly acidic pH (a pH of 5.6 to 6.4).118-120 The weak acidic pH is due to various substances, such as free fatty acids secreted from sebaceous glands or derived from phospholipid hydrolysis in the stratum corneum, lactic acid secreted from derived from the eccrine glands, urocanic acid mostly derived from the degradation products of filaggrin, and metabolites produced by microorganisms.121 The epidermis is also equipped with an active antimicrobial defense system comprising antimicrobial proteins (Table 14-3).
ANTIMICROBIAL PEPTIDES EXPRESSION FUNCTION
Human β-defensin 1 Epidermis Antimicrobial activity against gram-negative bacteria Role in keratinocyte differentiation
Human β-defensin 2 Epidermis Inducible by microbial components (Pseudomonas aeruginosa, Staphylococcus aureus, Candida albicans) Inducible by TNF-α IL-1 Antimicrobial activity against Escherichia coli and P. aeruginosa Weakly bacteriostatic against gram-positive bacteria
Human β-defensin 3 Epidermis Induced by TNF-α and bacteria Bactericidal activity against Gram positive bacteria (S. aureus and vancomycinresistant Enterococcus faecium)
Cathelicidin (LL-37) Keratinocytes Mast cells Neutrophils Eccrine gland ductal cells
Vitamin D is a major inducer Antibacterial, antifungal, and antiviral activities Attracts mast cells and neutrophils
Psoriasin (S100A7) Keratinocytes Bacterial membrane permeability Bactericidal against E. coli Chemoattractant for CD4 cells and neutrophils
RNase 7 Stratum
Broad spectrum antimicrobial
RNase 7 Stratum corneum Broad spectrum antimicrobial activity (S. aureus, Propionibacterium acnes, P. aeruginosa, E. coli, C. albicans) Induced by IL-1β, IFN-γ, bacterial challenge
activity (S. aureus, Propionibacterium acnes, P. aeruginosa, E. coli, C. albicans) Induced by IL-1β, IFN-γ, γ,γ bacterial challenge
corneum
IFN, interferon; IL, interleukin; TNF-α, tumor necrosis factor-α. From Modlin RL, Miller LS, Bangert C, et al. Innate and adaptive immunity in the skin. In: Goldsmith LA, Katz SI, Gilchrest BA, et al, eds. Fitzpatrick’s Dermatology in General Medicine, 8th ed. New York: McGraw-Hill; 2012.
222
ANTIMICROBIAL PROTEINS ON THE SKIN SURFACE
ANTIMICROBIAL PROTEINS
ON THE SKIN SURFACE
Antimicrobial proteins are evolutionarily ancient innate immune effectors produced by almost all plants and animals.122,123 Antimicrobial proteins show a broad antibacterial activity against both gram-positive and -negative bacteria; some even show antifungal or antiviral activity. The antimicrobial activity of most proteins occurs as a result of their unique structural characteristics, which enable disruption of the microbial membrane while leaving host cell membranes intact. These proteins also act as “alarmins” to alert host cells to react to injuries and microbial invasions.124
Many proteins act as antimicrobial proteins/ alarmins in the skin, including short proteins, such as β-defensins, cathelicidins, dermcidin, psoriasin, neuropeptides, and chemokines, and larger proteins, such as lysozymes, elastase, complement, and the S100 proteins. The variety of these antimicrobial proteins may reflect the complexity and long history of the microbial challenges at the skin surface. The epithelial cells lining the surface of the intestine, respiratory tract, reproductive tract, and skin are the major source of antimicrobial proteins. Each epithelium has many characteristic antimicrobial proteins that are necessary for each microenvironment. Cathelicidins and β-defensins are the two major antimicrobial proteins of the skin and show a broad spectrum of antimicrobial activity in vivo.125
CELLS THAT PRODUCE ANTIMICROBIAL PROTEINS
CELLS THAT PRODUCE
ANTIMICROBIAL PROTEINS
Keratinocytes produce various antimicrobial proteins to defend the skin (Fig. 14-12). In particular, keratinocytes in the hair follicle constitutively produce cathelicidins and β-defensins at a higher level than the keratinocytes of the interfollicular epidermis, probably because the microenvironment of hair follicles facilitates bacterial colonization much more readily than that of the interfollicular epidermis. Secretory cells of the skin, such as those in the eccrine, apocrine, and sebaceous glands, secrete their own antimicrobial proteins and lipids and contribute to the antimicrobial activity of the skin surface. In the dermis, mast cells produce large amount of cathelicidins and store them within intracellular granules. After skin injuries, mast cells secrete cathelicidins to resist bacterial and viral infections.126 Additionally, commensal bacteria produce their own antimicrobial proteins. For example, Staphylococcus epidermidis, the dominant commensal bacterium of the skin microbiota, produces several antimicrobial proteins.127,128 These various antimicrobial proteins derived from host cells and microorganisms can modulate or regulate the composition of the skin microbiota.
2
Antimicrobial proteins produced by host cells and commensal bacteria
Host-derived AMP Staphylococcus epidermidis
BD1, BD2, BD3 and BD4 Cathelicidin RNase7 Psoriasin Lactoferrin
Cathelicidin
Mast cell
Sebocytes
BD1 and BD2 Histone H4 Psoriasin Cathelicidin
Commensal-derived AMP (eg, PSMγ or PSMδ)
Lysozyme SLPI Elafin αMSH Catestatin Calprotectin
Stratum corneum
Viable cell layers of the epidermis
Dermis
Eccrine gland
Cathelicidin Dermcidin BD1 and BD2
Eccrine epithelial cells
FUNCTIONS OF ANTIMICROBIAL PROTEINS
FUNCTIONS OF
ANTIMICROBIAL PROTEINS
Antimicrobial proteins have various mechanisms of action122,129,130; many target the cell wall or cell membrane structure of microorganisms. For example, defensins and cathelicidins are usually cationic and interact with the bacterial membrane surface through electrostatic interactions. Some defensins form pores in the bacterial membrane to disrupt membrane integrity and promote bacterial lysis. Cathelicidins bind to bacterial membranes and promote membrane insertion and disruption. The mechanisms of action of several antimicrobial proteins remain unclear. Increasing evidence indicates that some antimicrobial proteins modulate immune signaling through chemokine receptors and Toll-like receptors. Antimicrobial proteins not only stimulate chemokine and cytokine secretion from a variety of cell types but also use chemotactic activity to recruit leukocytes by direct or indirect mechanisms to modify the inflammatory response.122,131-133 The chemotactic activities of different types of antimicrobial proteins are distinct from one another, and each recruits different types of
cell. For example, LL-37, the human member of the cathelicidin family of antimicrobial peptides, recruits neutrophils, T cells, monocytes, and mast cells. LL-37 also induces keratinocytes through the mitochondrial antiviral signaling pathway to promote the production and secretion of interferon-β, which activates antiviral responses.134 Toll-like receptors are transmembrane receptors for pattern recognition and are activated by the conserved structural pattern of microbial molecules, such as lipopolysaccharides, flagella, endotoxins, RNA, and DNA.135 Toll-like receptor signaling is not restricted to such microbial molecules but can also be initiated by some antimicrobial proteins, allowing the immune cell responses to be modified.136
EPIDERMAL BARRIER OF NEWBORN SKIN
Full-term newborn babies have a well-developed and functional skin barrier at birth.137 Although infant skin has a thinner stratum corneum with low levels of natural moisturizing factors and greater transepidermal water loss compared with adult skin, transepidermal water loss in healthy full-term newborns at birth is equal to or lower than that of adult skin (Table 14-4).138-140
223
2
COMPOSITIONAL DIFFERENCES PRETERM NEWBORN SKIN FULL-TERM NEWBORN SKIN INFANT SKIN ADULT SKIN REFERENCE
Epidermal surface Lacking the stratum corneum (<24 wk of gestation) Lacking the vernix caseosa (<28 wk of gestation)
Natural moisturizing factors None or only in a few layers of the stratum corneum (<26 wk of gestation)
Coated by the vernix caseosa Thin stratum corneum Thick stratum corneum 137,140,171
High in the vernix caseosa Low in the stratum corneum High in the stratum corneum 139,172
Surface pH — High Low Low 173-175
Sebum — — Low (7-12 mo old) High 176
Stratum corneum water content — — High Low 139
Dermal collagen fiber density — — Low High 140
Functional Differences
Rate of water absorption — — High Low 139
Transepidermal water loss Very high to high (depending
Low High Low 139
Transepidermal water loss Very high to high (depending on gestation period) Low High Low 139
on gestation period)
The stratum corneum is developed in utero under exposure to amniotic fluid, while extensive water exposure disrupts the stratum corneum barrier in adult skin. One putative mechanism for stratum corneum barrier development in utero is the formation of the fetal biofilm vernix caseosa, which coats the entire skin during the third (last) trimester.141 The vernix caseosa is a complex mixture of water, proteins, and lipids; fetal corneocytes swollen with water are distributed throughout an amorphous mixture of lipids. The vernix protects the epidermis from amniotic fluid and facilitates formation of the stratum corneum beneath it. Vernix retention after birth, compared with vernix removal immediately after birth, has been reported to result in greater skin hydration and a lower skin surface pH at 24 hours after birth.141 Moreover, vernix contains antimicrobial agents, including lysozyme and lactoferrin and exhibits antifungal and antibacterial activities.142-144 This evidence implies value in retaining the vernix rather than removing it immediately after birth, as recommended by the World Health Organization.145
Unlike a full-term infant, a premature infant has a weak epidermal barrier and mechanically fragile dermis at birth. Babies born before 28 weeks of gestation lack the covering of the vernix. Newborns with very low birth weight have a greater risk of skin damage as a result of their insufficient skin barrier. Babies delivered at 25 weeks of gestation have only a few layers of the stratum corneum at birth and show transepidermal water loss of about 70 g/m2/hr.146 The transepidermal water loss at birth in preterm newborns gradually decreases to about 7 g/m2/hr until 35 weeks’ gestation, around which time the epidermal barrier becomes well developed in utero. The epidermal barrier continues to develop after birth, but the transepidermal water loss of extremely preterm infants is significantly higher than that of full-term infants even at 1 month after delivery.146
224
BARRIER AGAINST PHYSICAL STRESSES
PHYSICAL STRUCTURE PROTECTING AGAINST MECHANICAL STRESSES
PHYSICAL STRUCTURE
PROTECTING AGAINST
MECHANICAL STRESSES
The skin, which protects our body from various mechanical physical stimuli, consists of stripes of rigid and soft tissues, that is, the rigid stratum corneum, soft layers of keratinocytes, rigid collagenous tissue of the dermis, and soft cushion of the hypodermis (which is rich in adipocytes). Deficits in the physical strength, and defects in the border, of each stripe cause various diseases that lead to physical weakness, such as epidermolytic ichthyosis; simple, junctional, and dystrophic epidermolysis bullosa; and Ehlers-Danlos syndromes (see Chaps. 15, 47, 60, and 72).
BARRIERS AGAINST ULTRAVIOLET STRESSES
BARRIERS AGAINST
ULTRAVIOLET STRESSES
Reflection at the air–skin interface, absorption by trans-urocanic acid, and diffraction via keratin filaments aligned parallel to the skin surface limit the penetration of UV radiation into the stratum corneum. In the viable cell layer of the epidermis, melanin is the major factor that absorbs UV irradiation and protects genomic DNA from UV-induced damage. Although the upper layer cells of the epidermis may undergo greater DNA damage by UV irradiation than basal
layer cells because of their location in the epidermis, the cells are tightly controlled in a nonproliferative state and are continuously eliminated from the epidermis via cell turnover, which prohibits tumorigenesis. In the proliferative basal layer cells, the damage to genomic DNA induced by UV irradiation is immediately repaired by DNA repair enzymes (see Chap. 130).
SWEATING TO PROTECT AGAINST HEAT STRESSES
SWEATING TO PROTECT
AGAINST HEAT STRESSES
The skin is a protective organ against heat and cold stress and plays an important role in controlling body temperature. In humans, cooling is mainly achieved by the evaporation of water secreted from eccrine sweat glands. Thermoplegia tends to occur in patients with congenital and acquired anhidrosis or hypohidrosis. Human skin has the highest reported density of eccrine sweat glands among all mammals.147 Most mammals have few sweat glands but have dense hairs covering their body. Because secreted sweat does not
2
readily evaporate under fur, it is reasonable that hairy mammals do not use sweat to cool their body. In contrast, the furless skin of humans has a high density of eccrine sweat glands and enables effective cooling via water evaporation from the body surface. Interestingly, the density of eccrine glands and hair follicles are conversely determined by one transcription factor, En1, in the mouse footpad.148
VASCULATURE AND BLOOD FLOW CONTROL AGAINST HEAT STRESSES
VASCULATURE AND BLOOD
FLOW CONTROL AGAINST
HEAT STRESSES
Cooling and maintaining heat are, in part, dependent on the control of blood flow at the skin surface and countercurrent heat exchange between arteries and veins.149,150 When the body temperature rises, skin surface blood vessels are dilated, and the skin appears red. The blood is cooled down via sweat evaporation and radiationinduced cooling on the skin surface (Fig. 14-13). In cold
Blood flow control in the skin for cooling and heat maintenance
Overheated
Superficial vein
Deep vein
Artery
Cold environment
Superficial vein
Deep vein
Artery Countercurrent heat exchange
Precooling of arterial blood by countercurrent heat exchange
Cooling
Sweat evaporation
Radiation cooling
Minimizing the heat loss
225
2
air, the skin turns pale because blood vessels contract to decrease blood flow and minimize heat loss via radiation cooling. Therefore, the skin regulates blood flow at the skin surface to maintain homeostasis of the body temperature. In the skin, arteries and veins run in close juxtaposition. Vascular countercurrent heat exchange between arteries and veins has been proposed as the mechanism by which the core body temperature is maintained under conditions of extreme cold or heat in homeothermic animals. For example, countercurrent heat exchange between afferent arteries and efferent veins in the legs of ducks prevents heat loss from the footpads, allowing ducks to paddle in icy water. In mouse skin, arteries and veins are aligned in close juxtaposition. Loss of the arteriovenous alignment in genetically engineered mice results in greater heat loss at the skin surface because warmer arterial blood reaches the skin surface without precooling via countercurrent heat exchange against the venous blood returned from the skin surface. The core body temperature decreases because of cooler venous blood returning to the body core.151 These observations, together with measurements of the blood temperature in the arteries and veins of humans,150 indicate that humans also use vascular countercurrent heat exchange to maintain the core body temperature under extreme conditions (see Fig. 14-13).
CONCLUSIONS
Skin, which is directly exposed to the external environment, shows species-specific adaptations, resulting in an amazing diversity among vertebrates and mammals and even among the human race and parts of the human body. It is important that dermatologists understand how human skin adapts to the external environment and how these adaptations are affected by disease. Skin is a complex superorgan consisting of the ectoderm, mesoderm, and skin microbiota. An integrated understanding of human skin, its microbiota, and the external environment that skin is exposed to is necessary for further investigation of normal skin function and of the molecular pathogenesis of various skin diseases; this will ultimately improve human health.
ACKNOWLEDGMENTS
The authors acknowledge the contributions of Ehrhardt Proksch and Jens-Michael Jensen, authors of Chap. 47, “Skin as an Organ of Protection,” in the 8th edition of Fitzpatrick’s Dermatology in General Medicine.
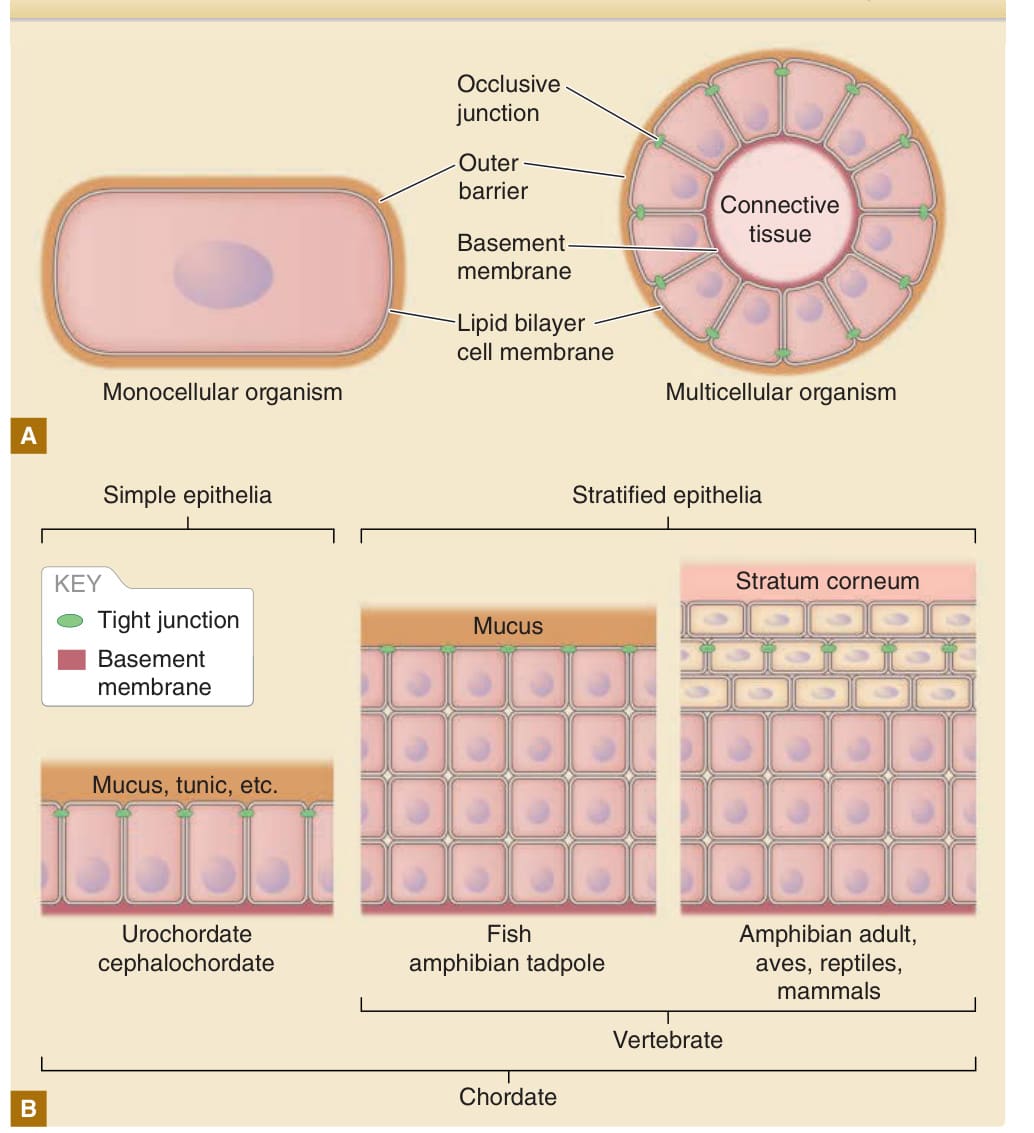
Figure 14-1 Basic structure of the surface barrier of unicellular and multicellular organisms. A, The lipid bilayer cell membrane is a basic diffusion barrier on the body surface of both monocellular organisms and multicellular organisms. In multicellular organisms, the intercellular spaces are sealed with occlusive junctions to limit leakage of the barrier through the paracellular pathway (see Fig. 14-9). In essence, both monocellular organisms and multicellular organisms are equipped with additional outer barriers (ie, a cell wall for monocellular organisms and mucus, cuticles, tunic matrix, or stratum corneum for multicellular organisms). B, Development of a stratified epithelium and stratum corneum were the two major evolutional events in vertebrate skin. Under the outer barrier, tight junctions seal the intercellular spaces in vertebrates.
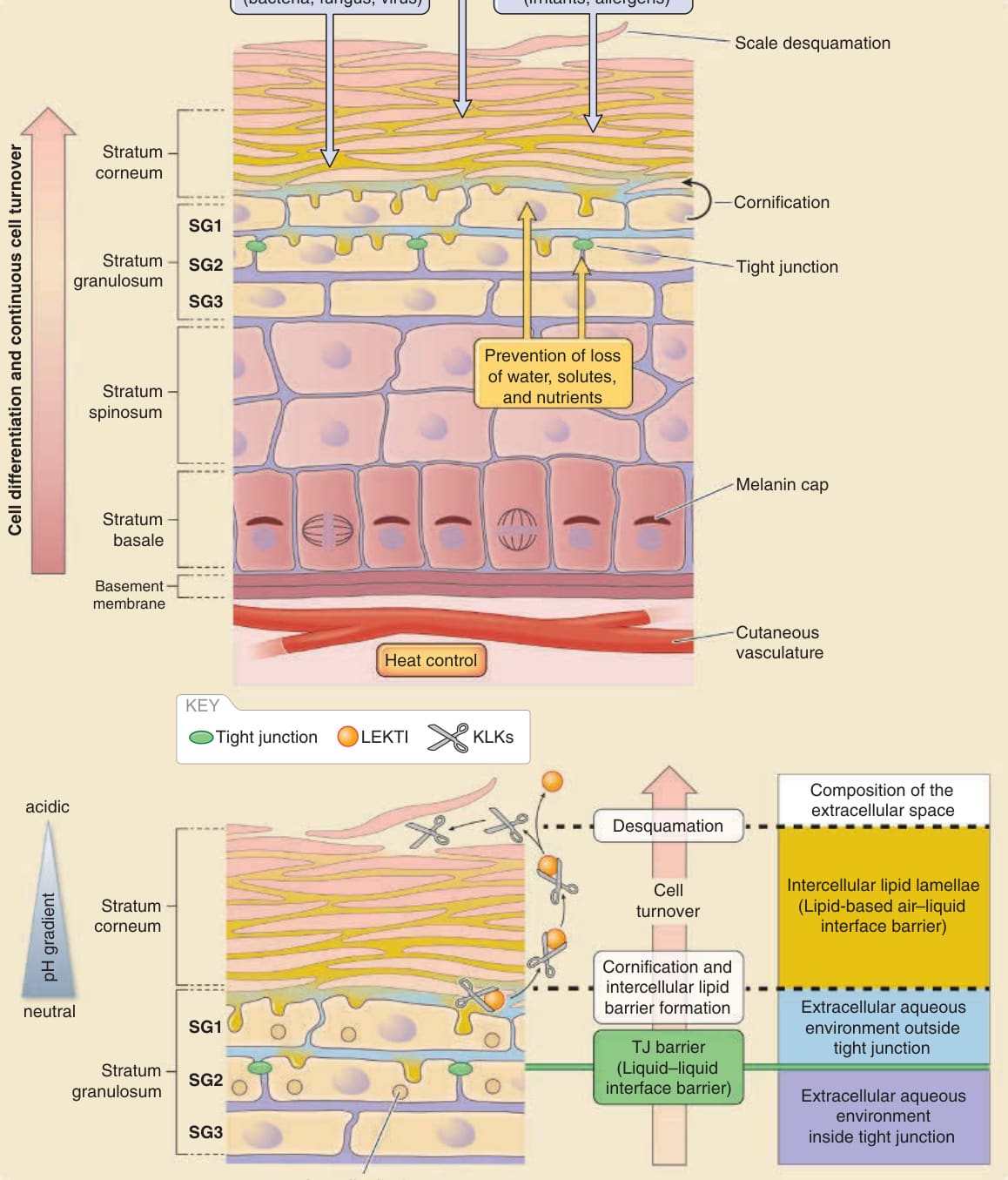
Figure 14-2 Basic structure of the epidermis and major epidermal barriers. A, Epidermal barriers protect the body against various outside-in physical, chemical, and microbial assaults in addition to inside-out leakage of water and solutes. Melanin caps protect the genomic DNA of basal cells from ultraviolet (UV) damage. Cells are continuously turned over to renew the epidermis and its barriers. At least three cell layers (SG1, SG2, and SG3 from outside to inside, respectively) exist in the stratum granulosum under the stratum corneum, where tight junctions (TJs) seal the intercellular spaces between the SG2 cells. B, Intercellular spaces are filled with lipids in, and with water under, the stratum corneum. The extracellular water environment is considered to be divided into two by the TJ barrier (extracellular space outside TJ barrier, light blue; extracellular space inside TJ barrier, purple). SG1 cells are ready to cornify at the outside TJ barrier. Contents of lamellar bodies (lipids, antimicrobial proteins, proteases, and protease inhibitors) are basically exocytosed from the apical surface of the SG2 cells to the outside TJ barrier. Kallikrein proteases (KLKs) are activated via the lower-pH-induced detachment of lymphoepithelial Kazal-type-related inhibitor (LEKTI) in the upper layers of the stratum corneum for desquamation. (Modified from Proksch E, Jensen J-M. Skin as an organ of protection. In: Goldsmith LA, Katz SI, Gilchrest BA, et al, eds. Fitzpatrick’s Dermatology in General Medicine, 8th ed. New York: McGraw-Hill; 2012.)
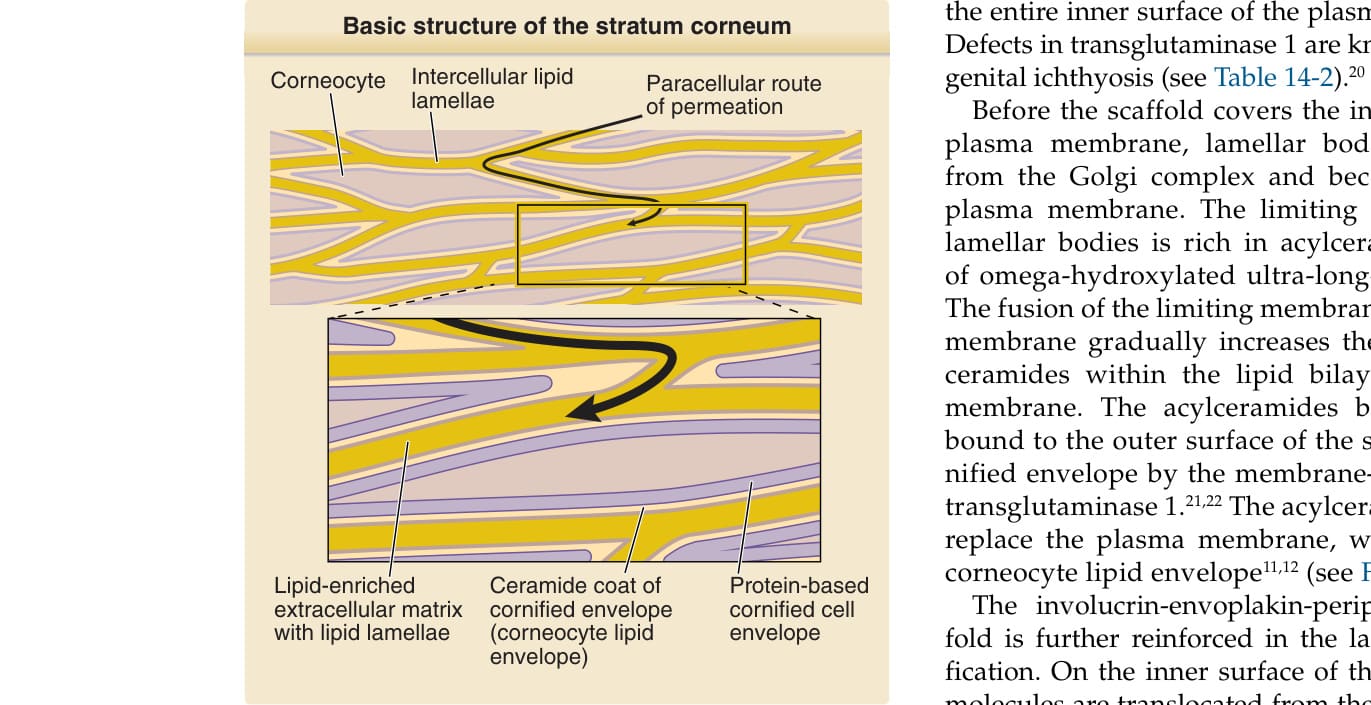
Figure 14-3 Basic structure of the stratum corneum. The stratum corneum comprises protein-based bricks (corneocytes) and lipid-based mortar (intercellular lipid lamellae). The surface of the corneocytes is coated with a thin layer of ceramide (lipid envelope). Details of how this structure is produced are shown in Fig. 14-6. (Modified from Proksch E, Jensen J-M. Skin as an organ of protection. In: Goldsmith LA, Katz SI, Gilchrest BA, et al, eds. Fitzpatrick’s Dermatology in General Medicine, 8th ed. New York: McGraw-Hill; 2012.)
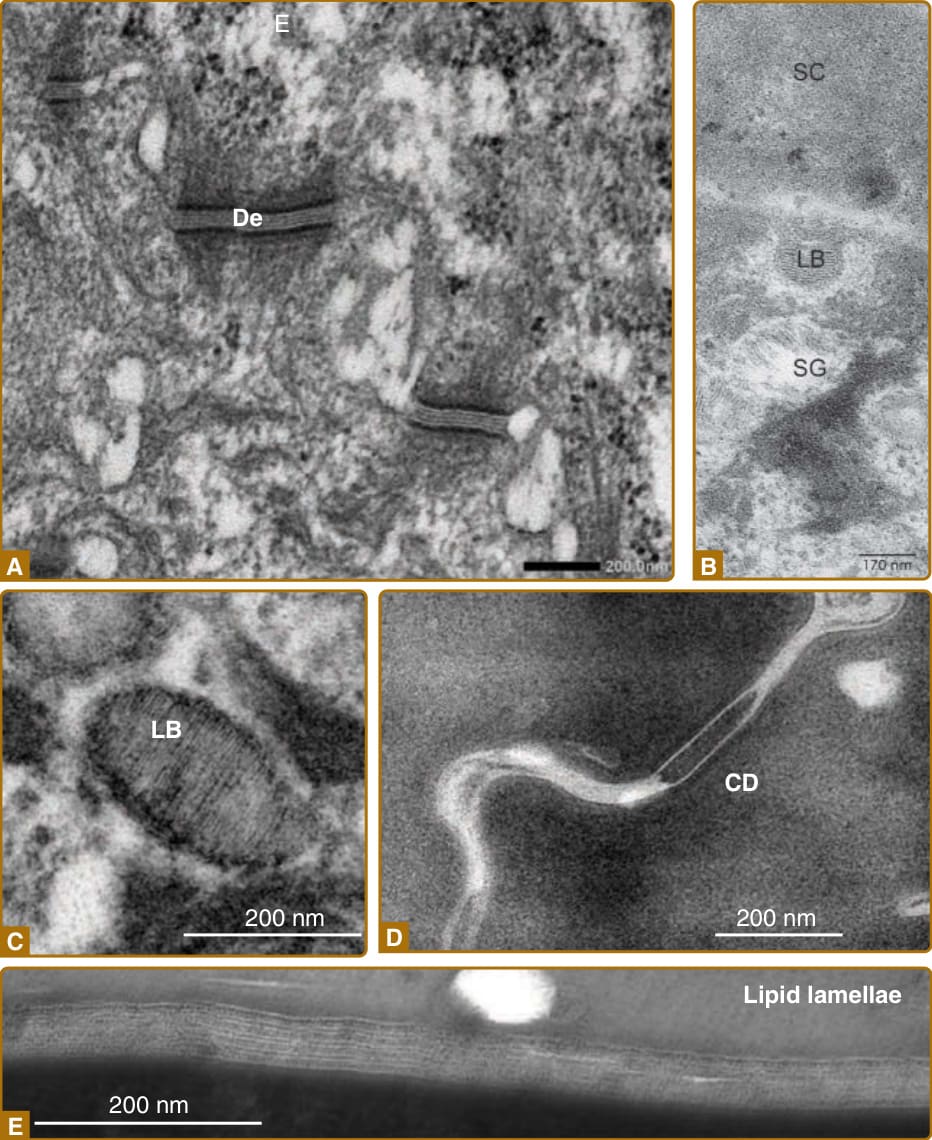
Figure 14-4 Major structures related to cornification, as seen under electron microscopy. A, Desmosomes (De) adhering to spinous layer cells. B, A lamellar body (LB) exocytosing its contents into the extracellular space at the apical surface of the SG1 cell. C, Contents of the lamellar body (LB) showing stripes of lipids. D, Corneodesmosomes (CD), cell–cell adhesion complexes between corneocytes. E, Intercellular lipid lamellae between corneocytes. (Images C and E, used with permission from Dr. Akemi Ishida-Yamamoto. Images A and D, used with permission from Dr. Toshihiro Nagai. Image B modified from Proksch E, Jensen J-M. Skin as an organ of protection. In: Goldsmith LA, Katz SI, Gilchrest BA, et al, eds. Fitzpatrick’s Dermatology in General Medicine, 8th ed. New York: McGraw-Hill;2012.)
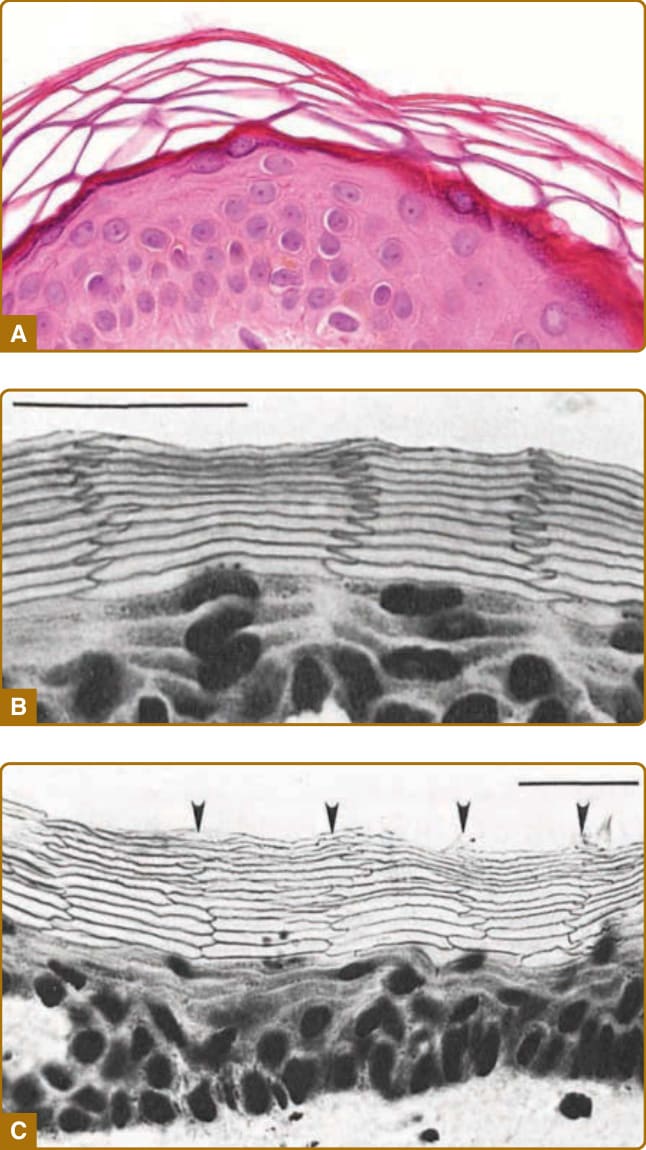
Figure 14-5 Morphology of the epidermis in cross-sections. A, Hematoxylin and eosin staining of paraffinembedded section of human skin showing the basket weave–like pattern of the stratum corneum. B, Frozen section of hamster ear skin stained with methylene blue, expanded in an alkaline solution with a pH of 12 and showing a regular alignment of corneocytes. A remarkably ordered structure of flattened stratum granulosum cells and anucleate corneocytes can be seen. The corneocytes show a regular zigzag interdigitation pattern between cell columns. C, Frozen section of human abdominal skin stained with methylene blue and expanded in an alkaline solution showing partially regular alignment of corneocytes. (Image B from Mackenzie IC. Ordered structure of the epidermis. J Invest Dermatol. 1975;65:45-51, with permission; Image C from Mackenzie IC, Zimmerman K, Peterson L. The pattern of cellular organization of human epidermis. J Invest Dermatol. 1981;76:459-461, with permission. Copyright © The Society for Investigative Dermatology.)
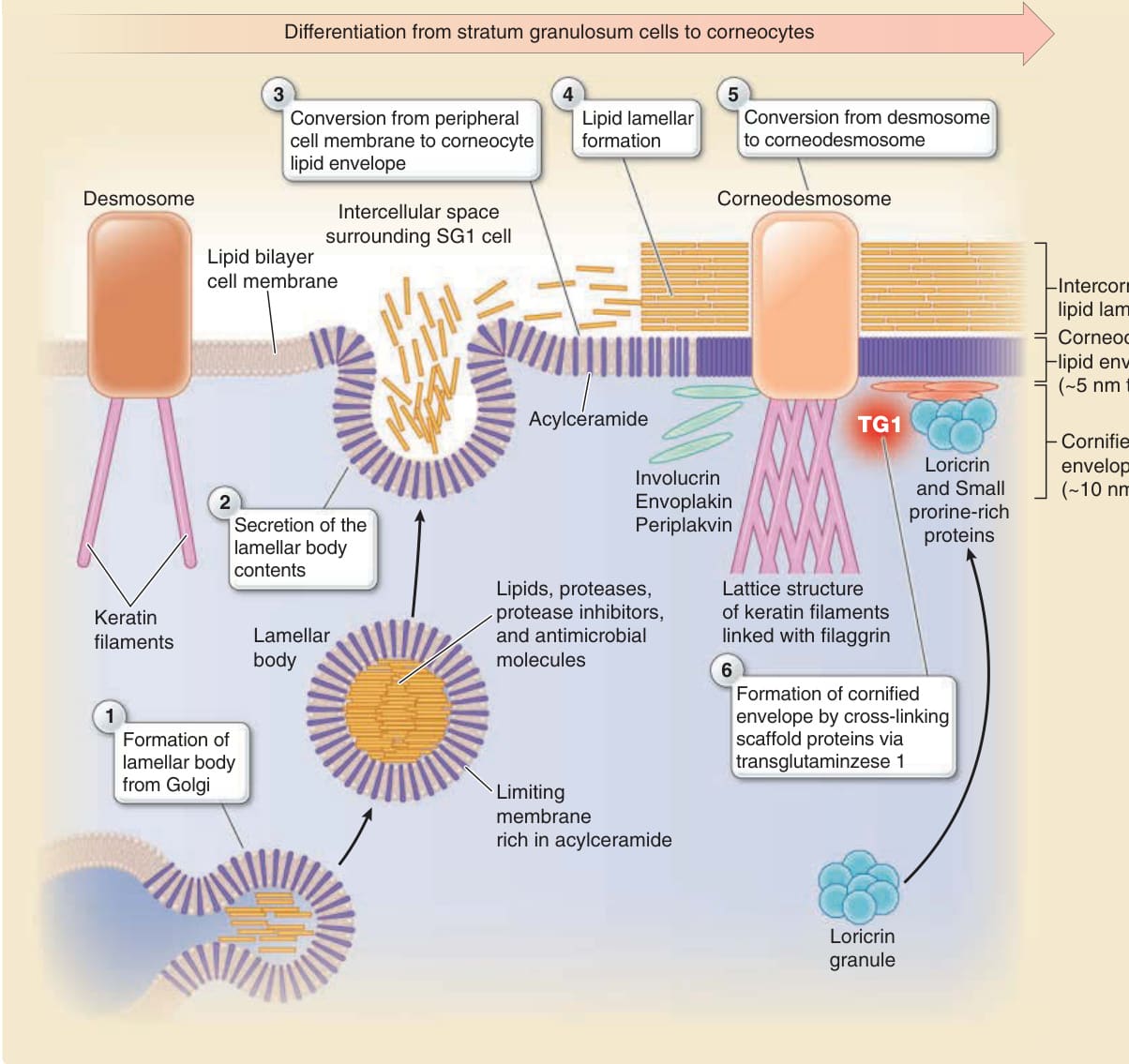
Figure 14-6 Formation of the cornified cell envelope, corneocyte lipid envelope, and intercorneocyte lipid lamellae. The time-dependent change of cornification (from SG2 cells via SG1 cells to corneocytes) is shown from left to right. A cornified cell envelope is formed beneath the lipid bilayer cell membrane via crosslinking of envoplakin, periplakin, and involucrin by transglutaminase 1 (TG1), which is further reinforced by loricrin in the later stage of cornification. Lamellar bodies exocytose their contents to the extracellular space outside tight junctions. The limiting membrane of the lamellar bodies contains a large number of acylceramides that replace the lipid bilayer cell membrane to form a corneocyte lipid envelope. As a result, the cornified cell envelope is coated by the corneocyte lipid envelope. The lipids exocytosed from lamellar bodies form the intercorneocyte lipid lamellae on the corneocyte lipid envelope to fill the intercellular space and form the water-resistant barrier of the stratum corneum. Corneodesmosin exocytosed from lamellar bodies is integrated into the desmosome, which is converted to the corneodesmosome. The lamellar bodies also exocytose proteases and protease inhibitors to control desquamation and antimicrobial proteins (see Fig. 14-12).
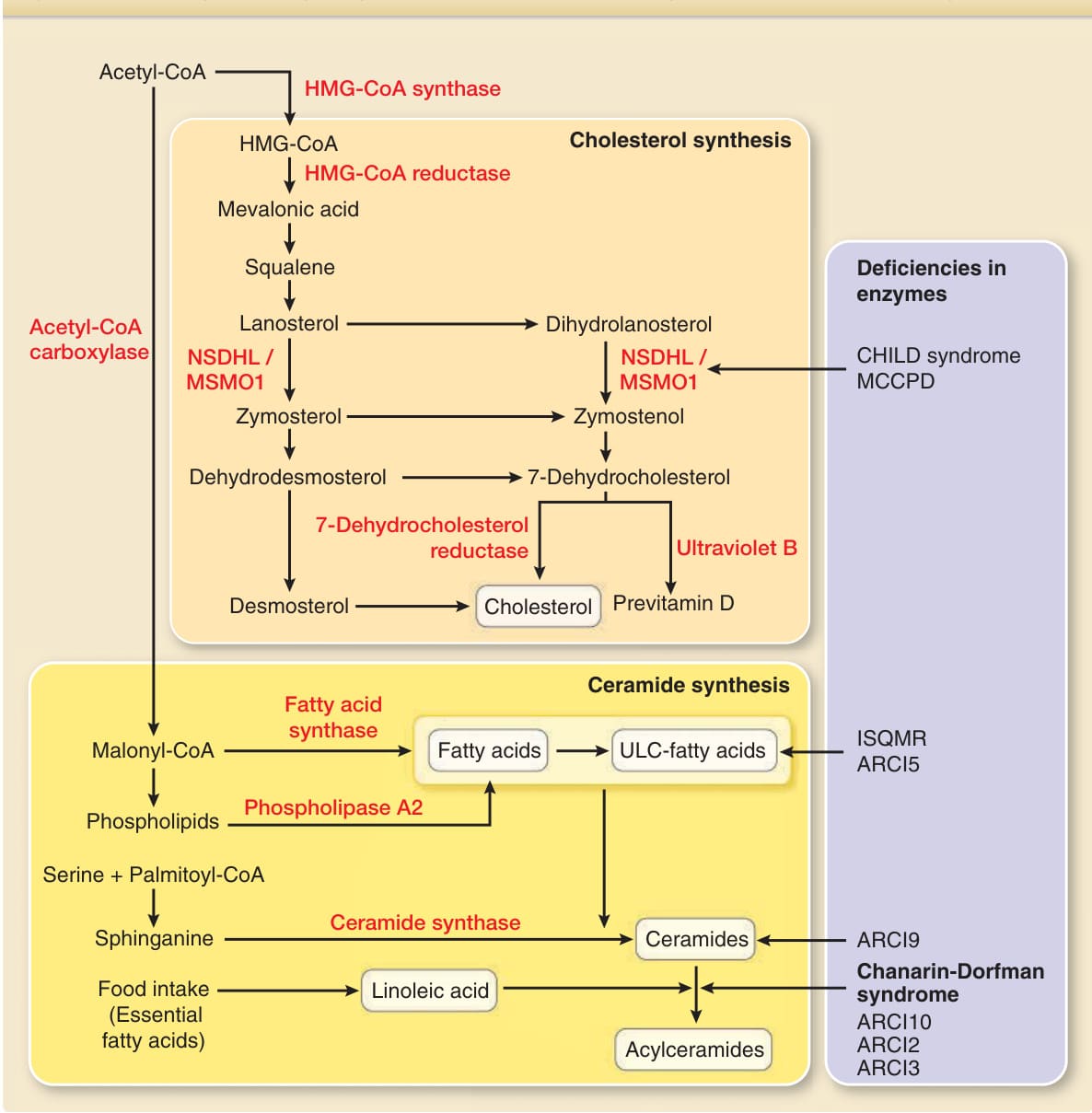
Figure 14-7 Synthetic pathways, key enzymes and deficiencies in the enzymes for cholesterol, free fatty acids, ceramides, and acylceramides. CoA, coenzyme A; HMG-CoA, hydroxymethylglutaryl CoA; CHILD syndrome, congenital hemidysplasia with ichthyosiform erythroderma and limb defects syndrome; MCCPD, microcephaly, congenital cataract, and psoriasiform dermatitis; ISQMR, ichthyosis, spastic quadriplegia, and mental retardation; ARCI, autosomal recessive congenital ichthyosis; NSDHL, NAD(P) dependent steroid dehydrogenase-like; MSMO1, methylsterol monooxygenase 1; ULC, ultra-long-chain.
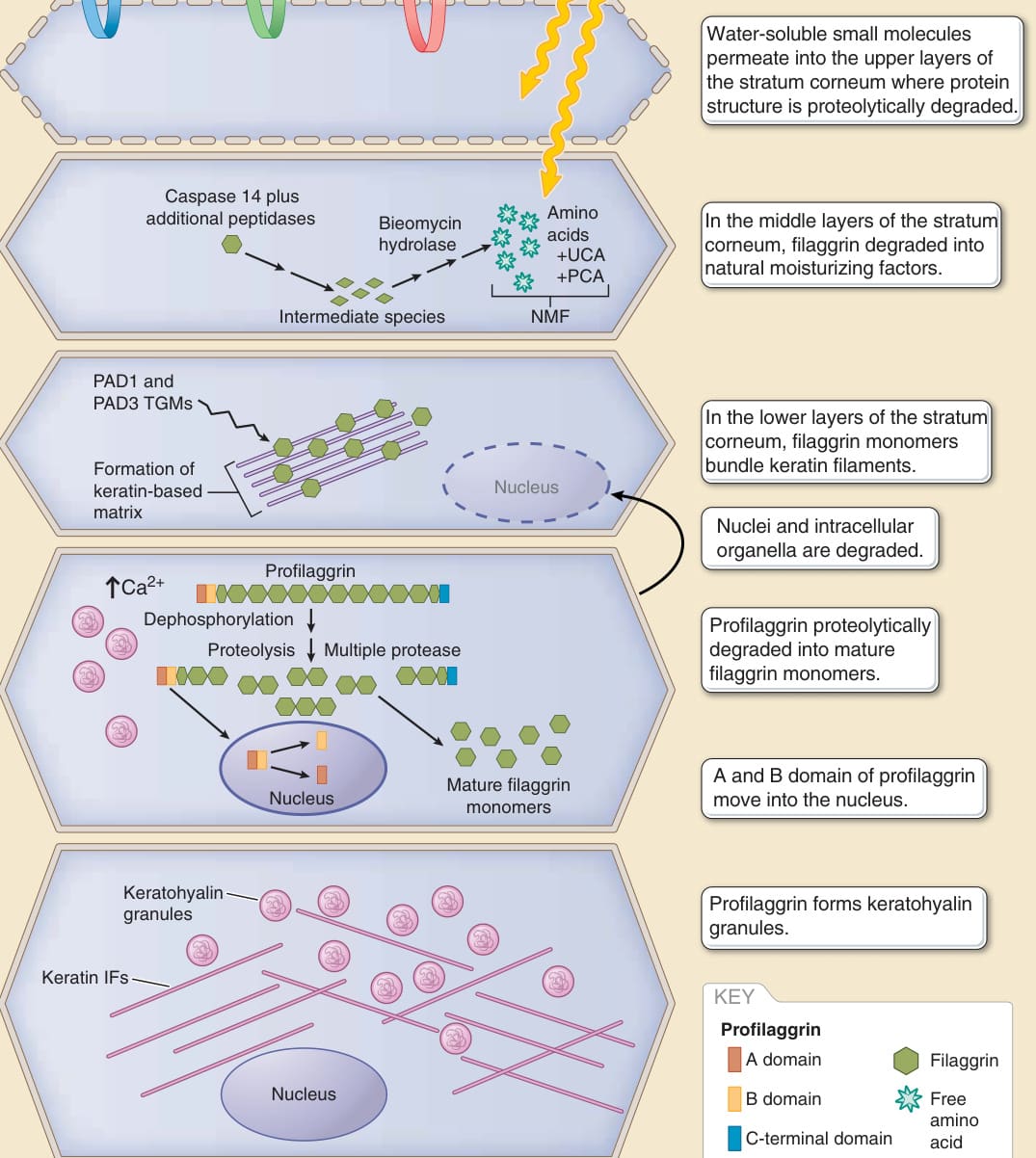
Figure 14-8 Multidomain structure of the stratum corneum and multistep maturation and degradation of filaggrin. The stratum corneum is thought to consist of at least three types of corneocytes undergoing different differentiation steps (corneocytes of the lower, middle, and upper stratum corneum). NMF, natural moisturizing factor; PAD, peptidylarginine deiminase; PCA, 2-pyrrolidone-5-carboxylic acid; IF, intermediate filament; SC, stratum corneum; TGM, transglutaminase; UCA, urocanic acid. (Modified from McAleer MA, Irvine AD. The multifunctional role of filaggrin in allergic skin disease. J Allergy Clin Immunol. 2013;131(2):280-291.)
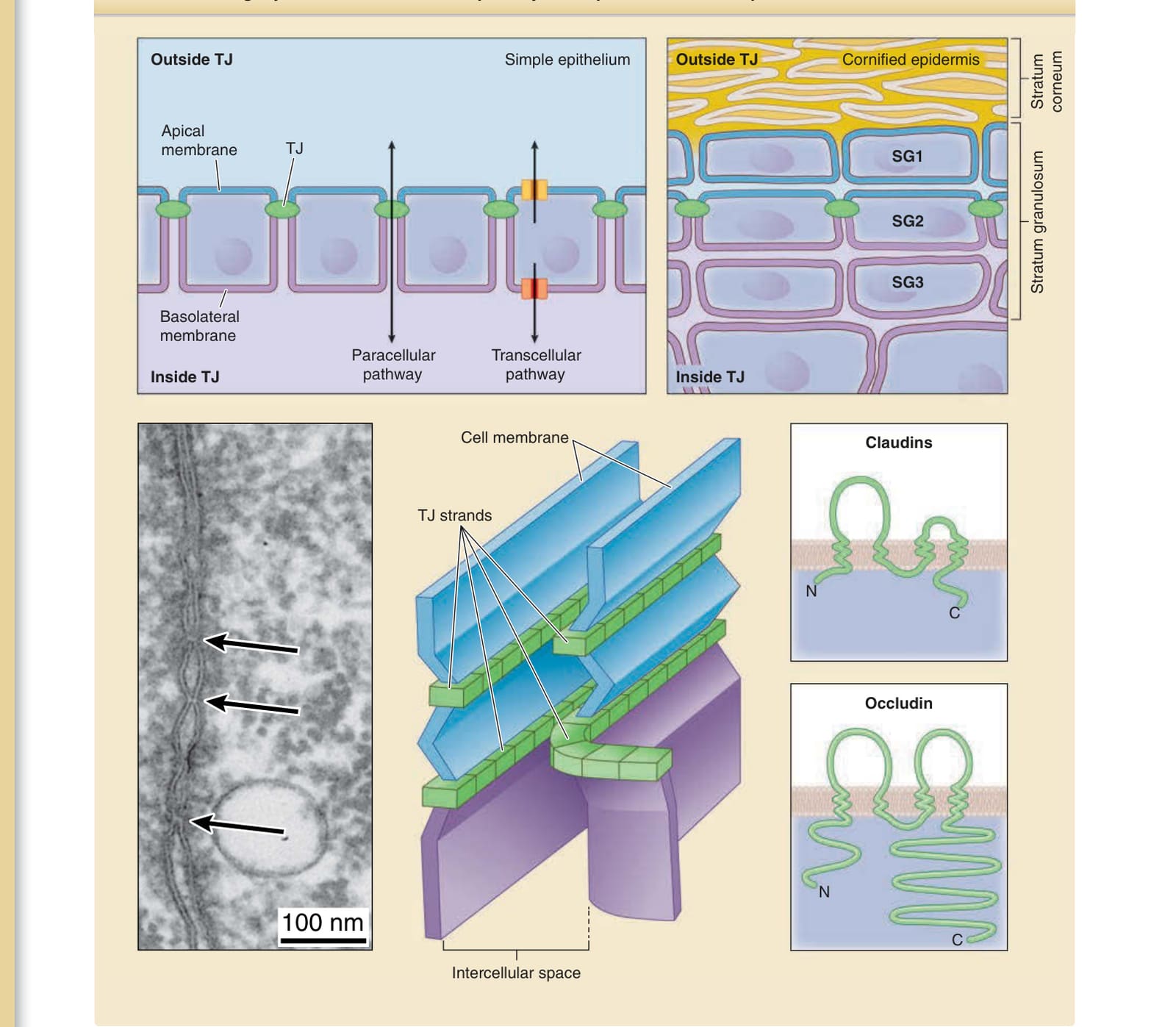
Figure 14-9 Tight junction (TJ) barrier in simple and stratified epithelial cellular sheets. The extracellular environment and plasma membrane are divided into two parts at the TJ barrier in simple epithelia. Solutes move between the two compartments via paracellular and transcellular pathways. In the epidermis, increasing evidence suggests that both the extracellular environment and plasma membrane are also divided into two parts at the TJ barrier formed between the SG2 cells, the extracellular environment outside and inside the TJ barrier, and the apical and basolateral cell membrane. In the electron micrograph (left panel), a TJ appears as a “kissing point” (arrows) where two plasma cell membranes face each other. The schematic shows the structure of TJ strands on the cell membrane and major transmembrane proteins of TJs. C, C-terminus; N, N-terminus. (Image used with permission from Dr. Hiroyuki Sasaki.)
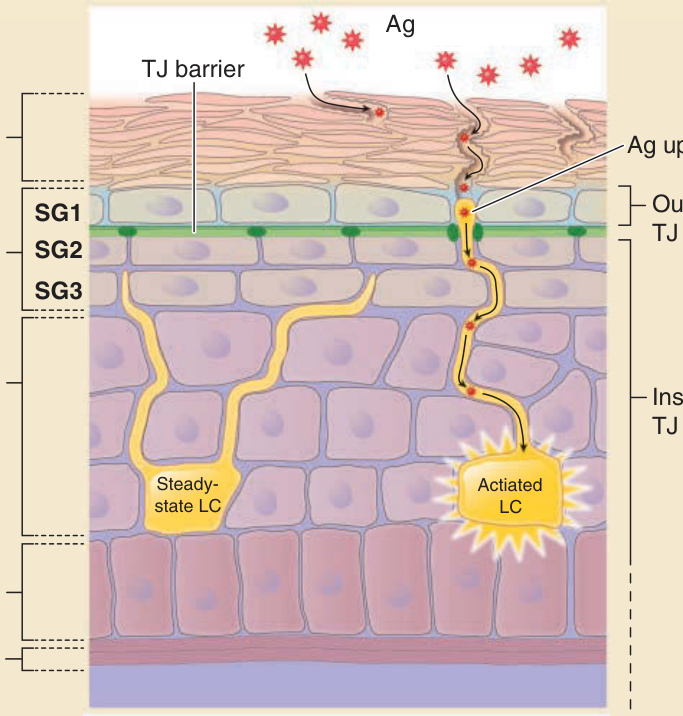
Figure 14-10 Spatial location of the stratum corneum, tight junctions (TJs), and Langerhans cells. SG2 cells form TJs (closed circle in dark green). SG1 cells are located outside the TJ barrier (light blue). In contrast, SG3 cells, spinous layer cells, and Langerhans cells are located inside the TJ barrier (purple). Dendrites of Langerhans cells in a steady state (left yellow cell) are located beneath the TJ but penetrate through the TJ barrier in their activated state (right yellow cell) to access to the outside TJ barrier. Antigen (Ag) uptake occurs from the tip of the dendrites (indicated by Ag uptake) at the outside TJ barrier.
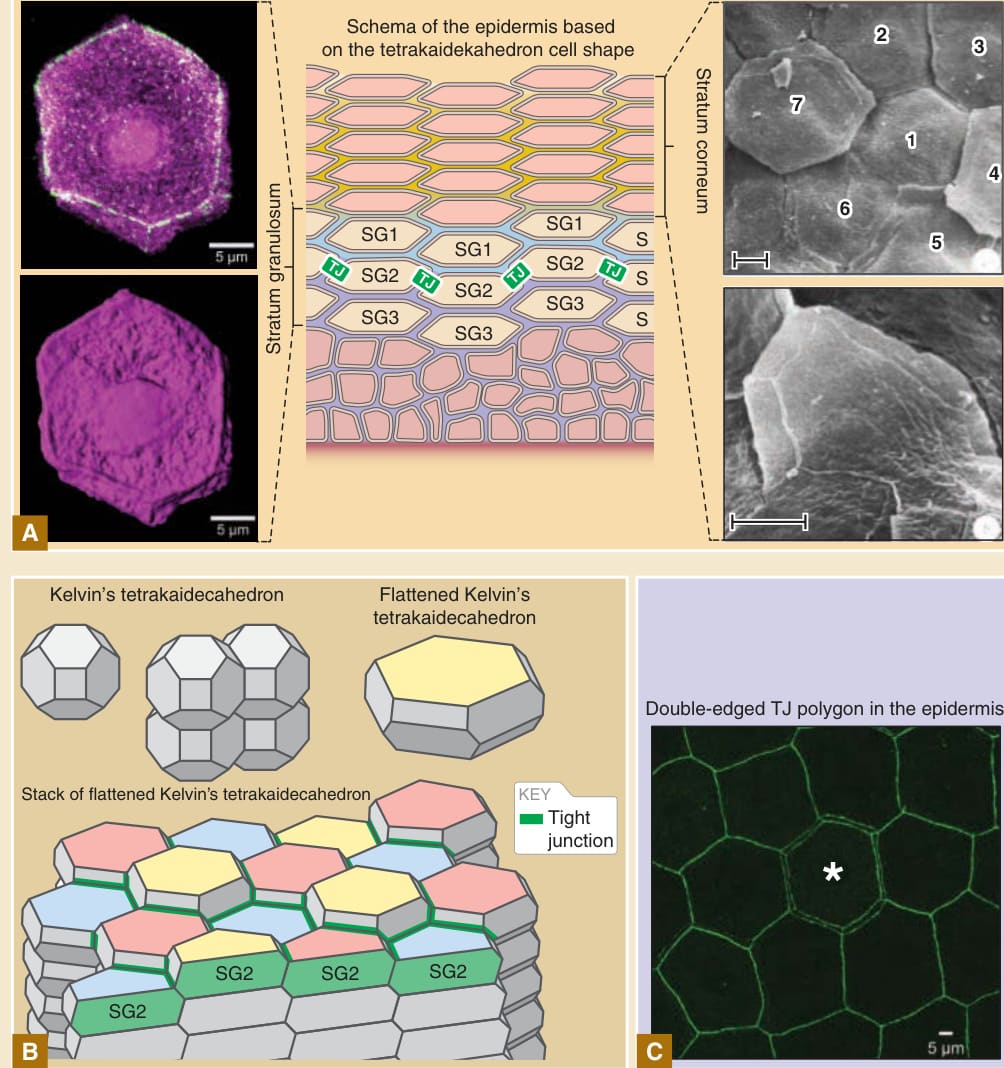
Figure 14-11 Three-dimensional cell shape of corneocytes and tight junction (TJ)–forming cells. A, Three-dimensional polyhedral shape of TJ-forming cells in immunofluorescent microscopic view (left panel) and the equivalent shape of corneocytes at the surface of the stratum corneum in scanning electron microscopic view (right panel) suggest that the basic shape of stratum granulosum cells and corneocytes is a flattened variation of Kelvin’s tetrakaidecahedron. B, Kelvin’s tetrakaidecahedron flattened variation of Kelvin’s tetrakaidecahedron, and regular interdigitated stacks of flattened Kelvin’s tetrakaidecahedron cells. SG2 cells are displayed at the top of cell columns. Three different Z-axis positions of the SG2 cells are displayed in yellow, pink, and blue on the apical surface of the cells. Tight junctions (green edges) located at the apical edges of the cell-cell contact between SG2 cells. C, En face view of the TJs (green edges stained by anti-ZO-1 antibody) in mouse ear skin. Essentially, tight junction polygons are single edged. The double-edged polygon (asterisk) is the site where a new TJ polygon is formed to relocate a cell from inside to outside of the TJ barrier. D, Conceptual model for the formation of the double-edged TJ polygon (asterisk) during cell turnover from inside to outside of the TJ barrier. A new TJ polygon is formed beneath the center yellow-colored cell (asterisk, left panel), resulting in the formation of a doubleedged polygon (middle panel). Then, the outer old polygon is degraded (right panel). The center yellow-colored cell in the left panel (asterisk) is relocated from inside the TJ barrier (left panel) to outside of the TJ barrier (right panel) after the temporal formation of the double-edged polygon (middle panel). (The scanning electron microscopic picture on the right side of Image A, is reproduced with permission of Springer Nature from Allen TD, Potten CS. Significance of cell shape in tissue architecture. Nature. 1976;264:545-547.) The immunofluorescent pictures of parts A and C and schemas in B and D are from Yokouchi M, Atsugi T, Logtestijn M, et al. Epidermal cell turnover across tight junctions based on Kelvin’s tetrakaidecahedron cell shape. Elife. 2016;5:e19593.
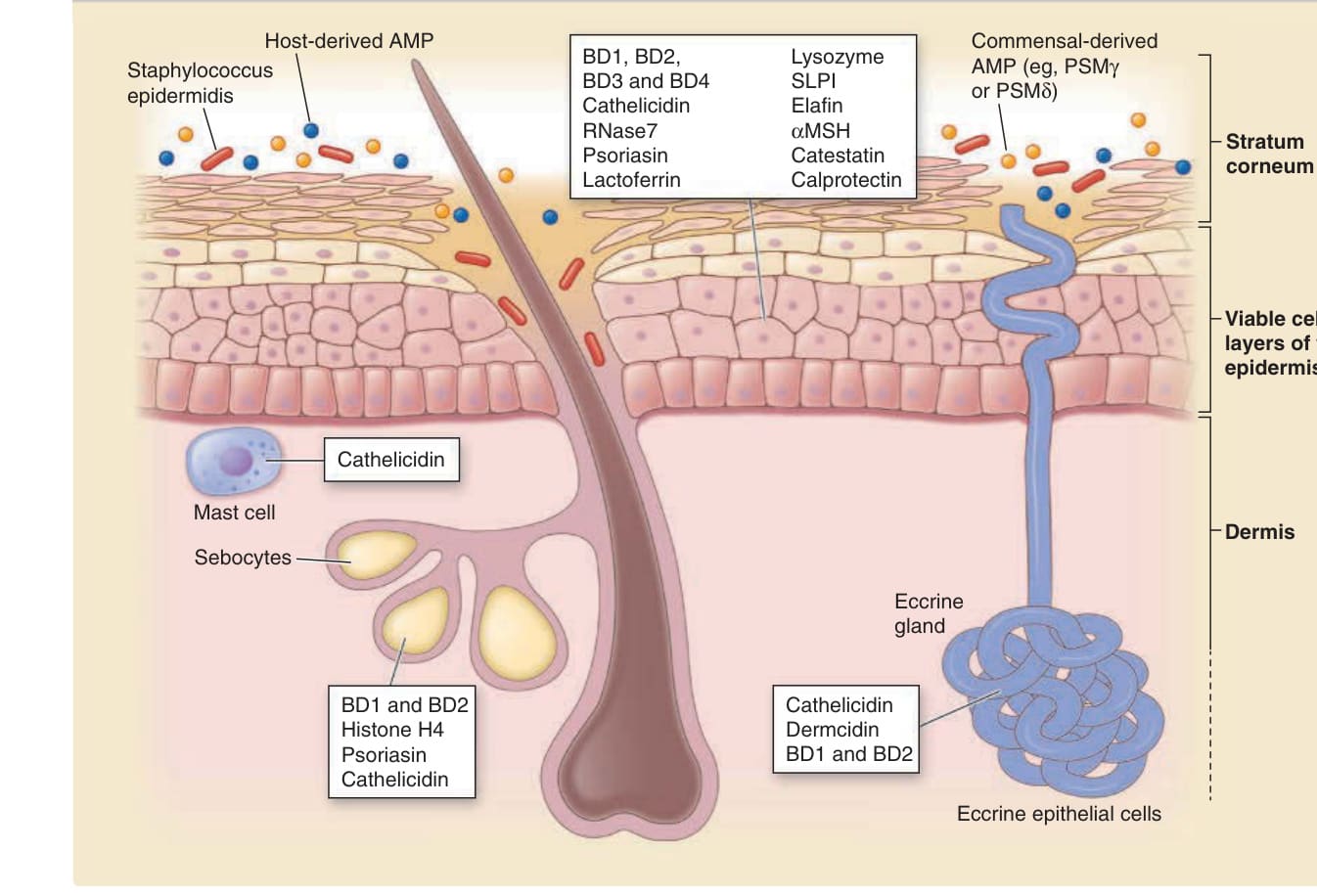
Figure 14-12 Antimicrobial proteins produced by host cells and commensal bacteria. AMP, antimicrobial proteins; BD, beta defencins; PSM, phenol soluble modulin; MSH, melanocyte-stimulating hormone; SLPI, secretory leukocyte protease inhibitor. (Adapted with permission from Springer Nature from Gallo RL, Hooper LV. Epithelial antimicrobial defence of the skin and intestine. Nat Rev Immunol. 2012;12:503.)
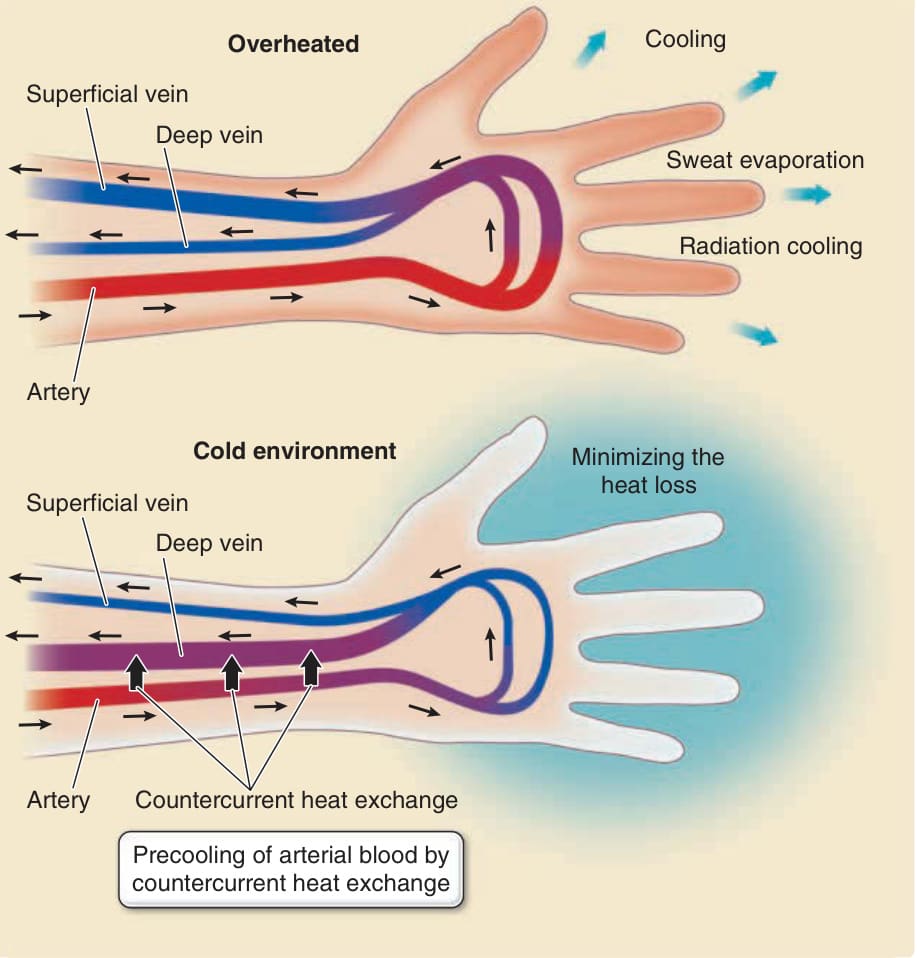
Figure 14-13 Blood flow control in the skin for cooling and heat maintenance. In a hot environment (upper panel), the superficial veins become dilated, and the blood in the superficial veins is mainly cooled via evaporation of eccrine sweat from the skin surface. In a cold environment (lower panel), the superficial veins contract, preventing heat loss from the blood. Countercurrent heat exchange between afferent arteries and dilated efferent deep veins precools the blood in the arteries before it reaches the superficial blood vessels, which also prevents heat loss from the skin surface.

TABLE 14-1 Short Commentary on Jargons
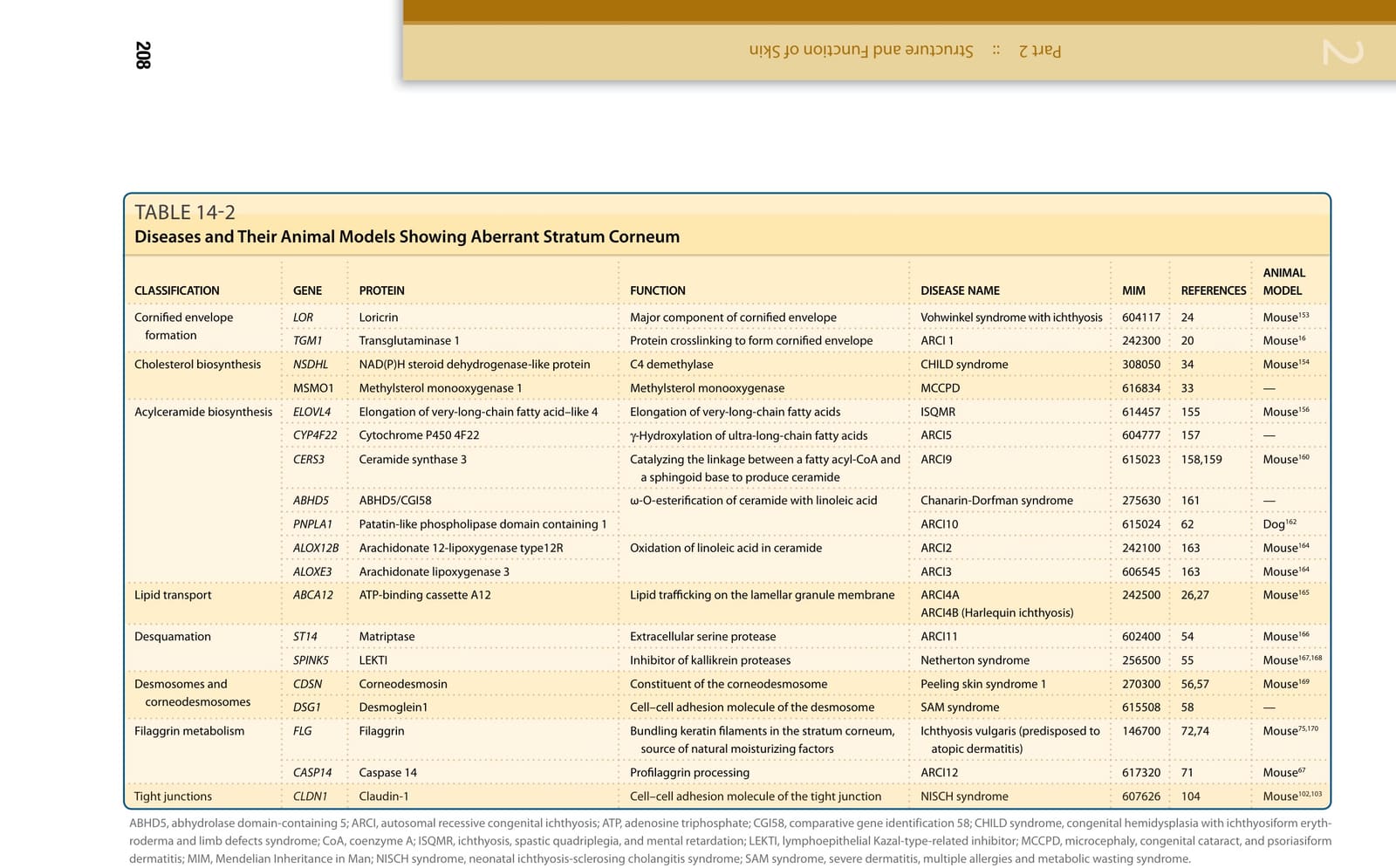
TABLE 14-2
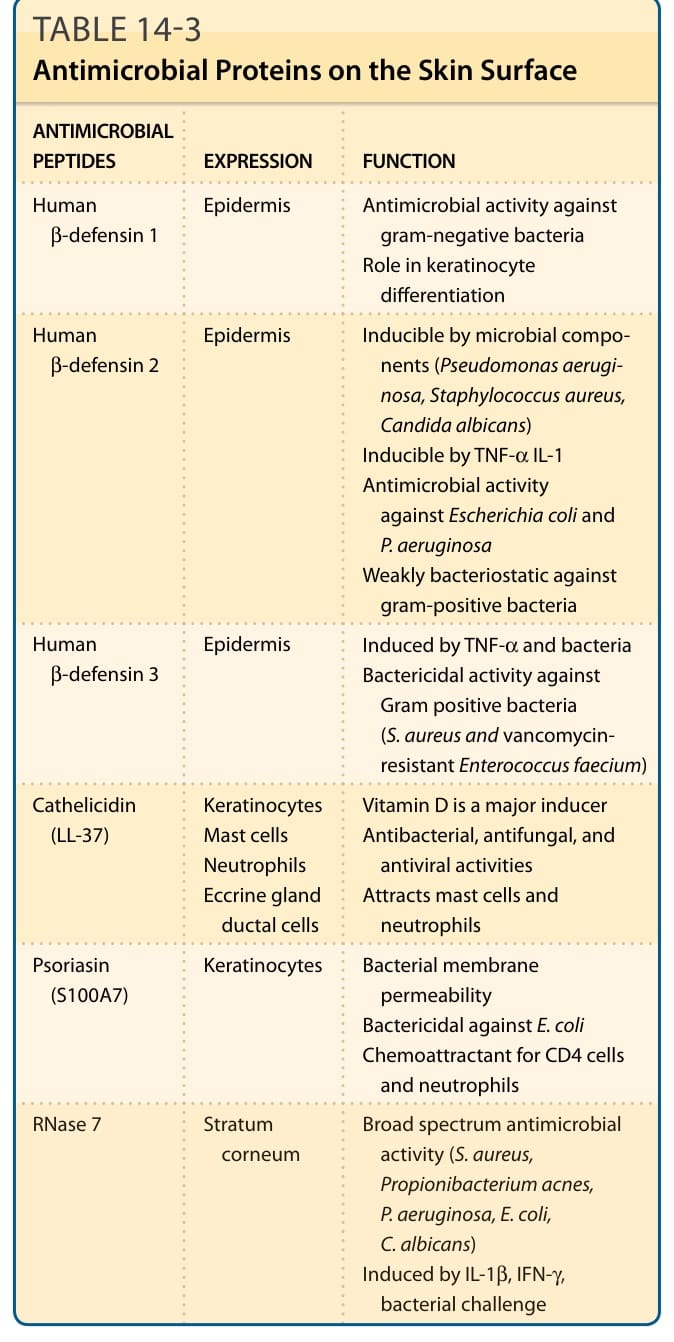
TABLE 14-3 Antimicrobial Proteins on the Skin Surface
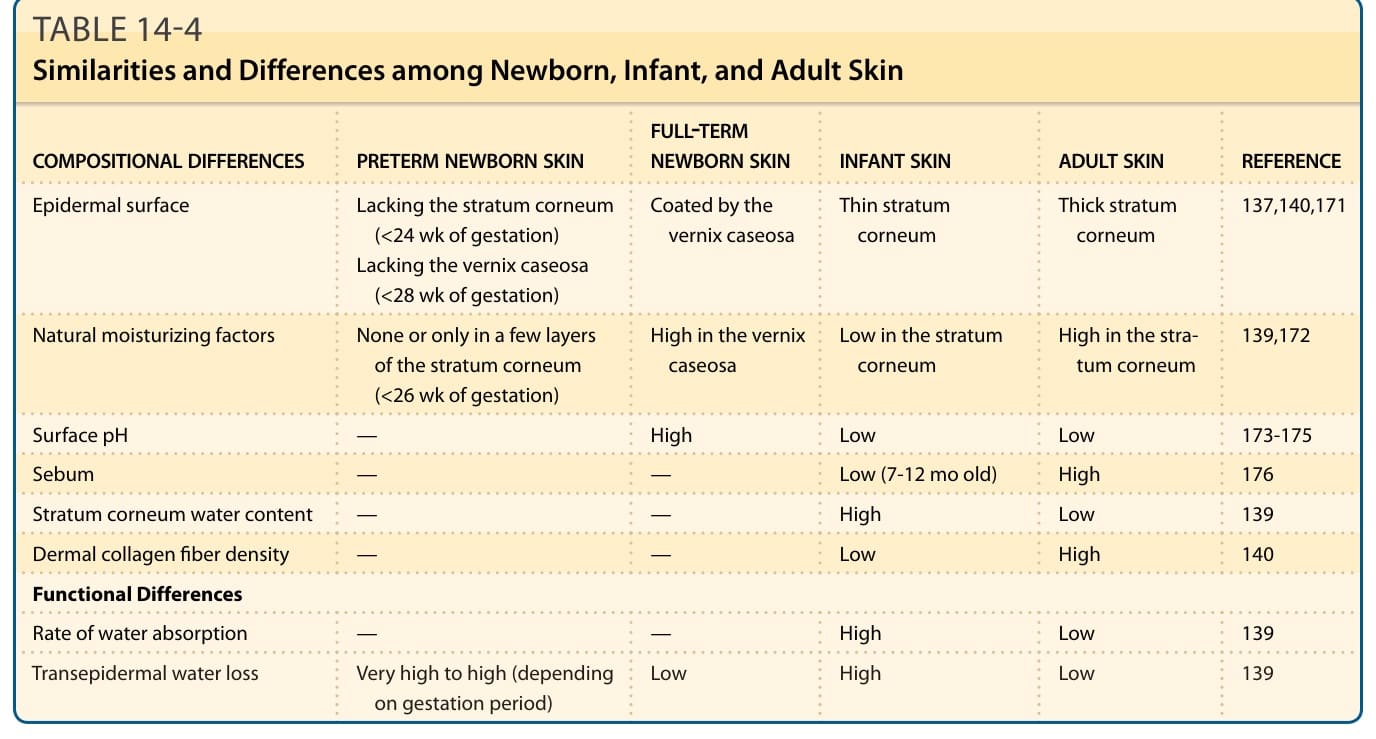
TABLE 14-4 Similarities and Differences among Newborn, Infant, and Adult Skin