指甲 (Nail) 精華筆記
甲器官構造與功能 (Anatomy and Function)
- 甲器官 (nail organ) 是皮膚附屬器 (skin appendage),位於每根手指與腳趾的遠端指(趾)骨 (distal phalanx) 上;生理狀態下持續生長,移除後可完全再生。
- 由近端往遠端,甲附屬器各層結構:
- 甲上皮 (eponychium):位於甲近端褶背側界線,在皮膚表皮與甲器官間形成保護性封閉。
- 甲近端褶 (nail proximal fold, NPF):皮膚表皮在甲上皮界線處向腹側內折、轉變為甲表皮後形成;是慢循環雙功能甲近端褶幹細胞 (NPFSCs) 的所在位置。
- 甲基質 (matrix):近端褶向背側遠端彎折後的腹側延續,由活躍增生的甲母細胞 (onychocytes) 組成;近端甲基質含快速增生的甲幹細胞 (NSCs)。
- 角化生成區 (keratogenous zone, KZ):甲基質上方,基質細胞在此分化、扁平化、死亡,沉積到甲板。
- 下甲皮 (hyponychium):甲上皮最遠端,在甲皮帶 (onychodermal band) 下方將甲板封閉至甲床,防止感染。
- 甲板 (nail plate):堅硬角化結構,由扁平無核的角質細胞 (corneocytes) 組成(由甲母細胞分化而來);保護指(趾)尖免於外傷、作為拾取小物的工具、施加反向壓力以增強敏感度。
- 正常表皮分化在近端褶內折後停止,故甲層不形成皮膚表皮的顆粒層 (granular layer)。
- 甲半月 (lunula,「小月亮」):透過甲板可見的白色新月形基底,是甲基質的可見部分;拇指最明顯,小指可能看不見。
- 甲床 (nail bed):甲基質遠端,由基底層與一至兩層基底上方有絲分裂後角質細胞組成,貢獻少數角化細胞至遠端甲板下表面;甲板與甲床交錯嵌合而附著。
- 甲角皮 (cuticle):由近端褶邊緣延伸至甲板,封閉甲近端、防毒素與外來物。兩側為外側甲溝 (lateral nail grooves) 與外側甲褶 (lateral nail folds)。
- 人類指甲較扁平;哺乳動物的爪呈近圓錐形、曲度大,形狀受下方遠端指(趾)骨與甲基質分布影響。
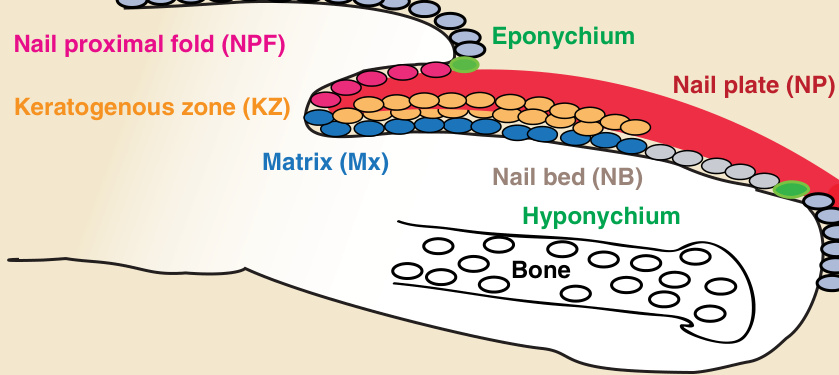
圖 8-2:人類指甲結構縱切面示意圖,標示各甲層與結構。
甲生長與分化 (Growth and Differentiation)
- 早期 Lewis 觀察甲板由三層組成,提出背側甲淺層源自甲近端褶、中間甲源自甲基質、深部腹側甲源自甲床。
- Zaias 與 Alvarez 以放射性氚標記甘胺酸 (tritiated glycine) 追蹤松鼠猴甲細胞,證明標記由甲基質移至甲板;甲床攝取極少而被排除。
- Berker 與 Angus 證明甲基質細胞高度增生、甲床相對不活躍。亦有研究依甲厚度與質量提出甲床對腹側甲板有貢獻。
- 目前共識:甲板主要由甲基質細胞所生成。
甲器官中兩個幹細胞庫 (Two Stem Cell Pools)
- 成體幹細胞 (adult stem cells) 需長期自我更新,分裂頻率低(避免突變併入),駐留於特化微環境 (niches) 中。活化後離開微環境形成快速增生的過渡放大型 (transit-amplifying, TA) 前驅細胞。
- 慢循環特性可用於辨識幹細胞:脈衝追蹤法以 ³H-thymidine、溴去氧尿苷 (bromodeoxyuridine, BrdU)、或四環素誘導型 H2B-GFP 轉殖小鼠標記,慢循環細胞保留標記成為標記保留細胞 (label-retaining cells, LRCs)。Cotsarelis 等以此在毛囊隆突區 (bulge) 與角膜緣 (cornea limbus) 辨識出 LRCs。
- 慢循環雙功能 NPFSCs:H2B-GFP 系統在 NPF 基底層辨識出甲 LRCs(小鼠中以環狀排列),表現毛囊幹細胞標記 K15、K19、PHLDA1,極少 Ki67⁺ 細胞而較靜止。
- 譜系追蹤顯示 K15 標記細胞同時貢獻甲結構與更主要的甲周表皮 (perinail epidermis),故為雙功能 (bifunctional)。
- BMP 訊息傳導為正確甲形成與分化所必需:缺乏時甲基質增生減少、KZ 消失,甲採取表皮命運(顆粒層延伸、表現 K1 與 loricrin,而甲板標記 AE13 偵測不到);亦為維持 NPFSCs 之 LRC 特性所必需。Msx2、Foxn1 為 BMP 下游標靶。
- 快循環單能性 NSCs:甲基質為 NPF 形態學延續,經中間區 (intermediate zone, IZ) 相連。IZ 隨追蹤期縮小(2 週最小、4 週消失),顯示由 NPF 慢循環 LRCs 至甲基質快速分裂細胞的細胞分裂梯度。
- Takeo 等以 K14-Cre-ER 譜系追蹤證明 LacZ⁺ 條紋僅源自近端甲基質;近端甲基質細胞表現 K14、K17 與高度 Ki67,體外群落形成能力最高,符合 NSCs 標準。
- Wnt 訊息傳導直接參與甲分化:Wnt 訊號在近端甲基質缺失、向遠端甲基質開啟;移除 β-catenin 或 Wntless 後整個甲上皮呈 NSC 特性(K17⁺、Ki67⁺)且缺 AE13。
- Lgr6(富含白胺酸重複序列的 G 蛋白偶聯受體 6)為 NSCs 特有標記,定位於近端甲基質,產生甲板。
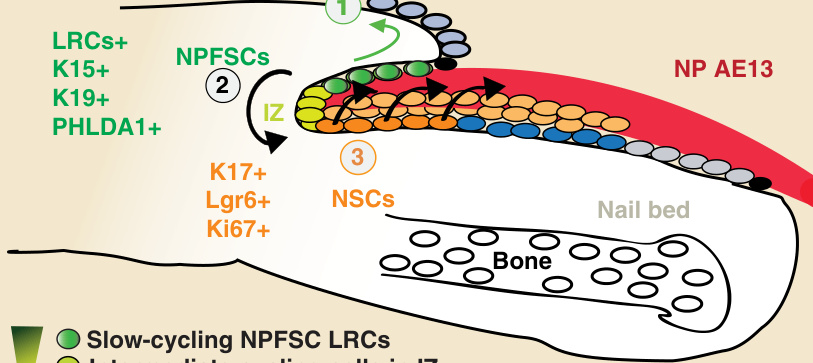
圖 8-4:指甲中兩個幹細胞族群(NPF 的 NPFSCs 與近端甲基質的 NSCs)定位示意圖。
甲上皮在指(趾)端再生中的角色 (Digit Regeneration)
- 人類與小鼠的指(趾)端能在受傷後完全再生,此能力僅限於與指甲相關的區域;中節指(趾)骨截肢後甲移植可誘導異位指(趾)骨分化。
- 近端甲基質的 NSCs 及其分化與協調指(趾)端再生的能力直接耦合;過程依賴上方覆蓋的甲器官與下方胚芽 (blastema) 的交互作用。
- 分子模型(小鼠):近端甲基質 NSCs 透過 Wnt 活化 (Wntless) 產生遠端甲基質細胞,兩者分化為甲板。遠端截肢後,再生甲上皮活化 Wnt → 經 semaphorin 5a (Sema5a,軸突導引分子) 促進胚芽神經支配 → 神經支配為再生甲上皮表現 FGF2 (fibroblast growth factor 2) 所必需 → FGF2 促進 Runx2 陽性前驅細胞或 Sp7 成骨細胞 (osteoblasts) 增生 → 耦合指(趾)端再生。FGFR1 表現於間質胚芽,可能招募 ERK 路徑。
- β-catenin 條件性剔除導致甲分化缺失(缺 TopGal、AE13 表現)、骨再生受阻、Sema5a 下調。去神經支配 (denervation) 抑制胚芽生長。
- 在 NSC 微環境近端截肢後,β-catenin 穩定化無法恢復指(趾)端再生(與遠端甲基質功能形成對比);消融 Lgr6 亦抑制再生。Lgr6 亦表現於骨成骨細胞與外分泌汗腺 (eccrine sweat glands),貢獻胚芽。
- 相關人類甲疾病:遺傳性無甲症 (inherited anonychia) 與孤立性先天性甲發育不良 (isolated congenital nail dysplasia);遺傳性無甲症家族有嚴重甲發育不全 (hypoplasia),辨識出與 Wnt 路徑相關的 R-spondin 4 (Rspo4) 突變。
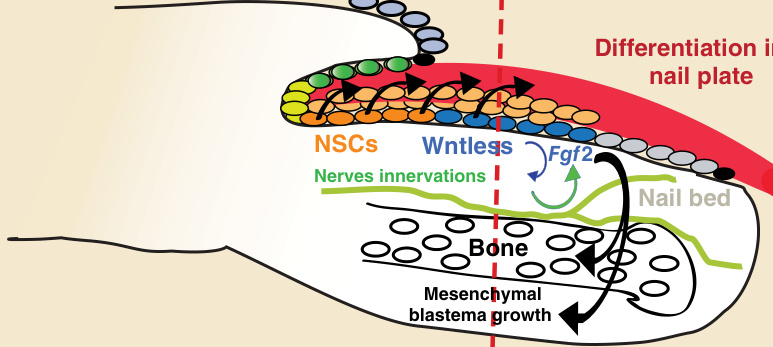
圖 8-6:甲上皮分化與指(趾)端再生的分子連結。
結論 (Conclusions)
- 甲器官含一個由慢循環到快循環幹細胞的梯度:近端褶慢循環 NPFSCs → IZ 較活躍細胞 → 近端甲基質快速增生 NSCs,類似毛囊幹細胞 (hfSCs) 的動態階層。
- 類比腸道兩個幹細胞族群(+4 位置慢循環 Bmi1⁺ LRCs 與隱窩底快循環 Lgr5⁺),推測指甲中 NPFSCs 與 NSCs 間可能存在備份系統——慢循環 NPFSCs 在壓力、損傷或幹細胞耗竭時作為 NSCs 的儲備。
- NSC 分化與指(趾)端再生有直接關係,NSCs 可能具治療潛能,未來或能協調完整指(趾)端再生(含骨、神經支配、血管與皮膚附屬器),造福截肢病人。