指甲 (Nail)
2
重點一覽 (AT-A-GLANCE)
甲附屬器 (nail appendage) 由數層結構所組成,這些結構共同建構出甲器官 (nail organ):
■ 甲上皮 (eponychium) 在甲近端褶 (nail proximal fold, NPF) 的背側界線處,於皮膚表皮 (skin epidermis) 與甲器官 (nail organ) 之間形成界線,構成一道保護性封閉。
■ 甲近端褶 (NPF) 是在皮膚表皮於甲上皮 (eponychium) 界線處向腹側內折、轉變為甲表皮 (nail epidermis) 之後形成的,由此構成 NPF。NPF 是慢循環、雙功能的甲近端褶幹細胞 (nail proximal fold stem cells, NPFSCs) 的所在位置。NPFSCs 在甲再生時,會主動將子代細胞輸送至甲周表皮 (perinail epidermis) 與甲基質 (nail matrix),並隨之輸送已分化的甲板 (nail plate)。
■ 甲基質 (matrix) 是近端褶在向背側、遠端彎折之後的腹側延續,由稱為甲母細胞 (onychocytes) 的活躍增生細胞所組成。在近端甲基質 (proximal nail matrix) 中,存在快速增生的甲幹細胞 (nail stem cells)。其分化能力與協調指(趾)端再生 (digit regeneration) 的能力直接耦合。
■ 甲基質分化後形成角化生成區 (keratogenous zone),最終將細胞沉積到其上方覆蓋的甲板中。
■ 下甲皮 (Hyponychium) 是甲上皮最遠端的部分,位於甲床 (nail bed) 的周邊、甲板下方,在游離緣 (free edge) 與指尖皮膚表皮交界處,形成一道封閉以保護甲床。
前言 (INTRODUCTION)
甲器官 (nail organ) 是人體皮膚附屬器之一,位於每一根手指與腳趾的遠端指(趾)骨 (distal phalanx) 上。有趣的是,指甲在生理狀態下會持續生長,並能在移除後完全再生。本章涵蓋甲附屬器生物學、其生長調控以及甲分化機制等方面的新發現。科學家一直密集地在各種器官中尋找幹細胞,期望了解並善用其再生能力。事實上,皮膚是一個複雜的器官,其中包含許多不同的「微型器官 (miniorgans)」——即由各自獨立幹細胞族群所維持的皮膚附屬器。過去數年來,學界對於甲器官本身是否擁有能夠再生整個甲附屬器的幹細胞,或是否有其他細胞參與甲再生,進行了積極的研究與討論。本章呈現有關甲器官中共存兩個獨立皮膚幹細胞族群(兩者具有相當相反的細胞週期動態)的新發現,亦即:慢循環的雙功能甲近端褶幹細胞 (slow-cycling bifunctional nail proximal fold stem cells, NPFSCs) 與快循環的甲幹細胞 (fast-cycling nail stem cells, NSCs)。此外,本章也討論了它們在甲器官再生過程中的相互依賴與交互作用。因此,指甲含有一個由慢循環到快循環細胞的梯度,從慢循環的 NPFCs,到中間區 (intermediate zone, IZ) 中較活躍的細胞,再到近端甲基質中高度增生的幹細胞,逐步將兩者分隔開來。我們也介紹甲器官的一項新功能,特別是新發現的近端甲基質中的 NSCs,以及其位於遠端甲基質中的相鄰前驅細胞,它們積極參與甲分化,並在哺乳動物指(趾)端再生的協調中扮演令人驚訝的關鍵角色。此外,從甲器官中分離幹細胞並進行其基因表現分析,揭示了它們的分子特徵,其中骨形態發生蛋白 (bone morphogenetic protein, BMP) 與無翅型 MMTV 整合位點家族 (wingless-type MMTV integration site family, Wnt) 訊息傳導路徑被確認為甲命運分化 (nail fate differentiation) 以及指(趾)端再生中的必要因子。因此,目前的研究已開始揭示包括指甲在內的其他皮膚附屬器也擁有各自具再生能力的幹細胞。然而,仍需更多研究以進一步描繪甲器官中這些幹細胞的特性,這可能在未來為人類指甲假設性的幹細胞再生潛能帶來一些啟發。因此,由於其非凡的再生潛能,進一步研究調控指甲幹細胞族群的機制及其與間質細胞 (mesenchymal cells) 的交互作用,可能會帶來治療各種甲疾病(包括人類截肢 amputations)的新途徑。
甲器官的構造(解剖)與功能 (NAIL ORGAN STRUCTURE [ANATOMY] AND FUNCTION)
指甲屬於皮膚附屬器之一,位於人體每一根手指與腳趾的遠端指(趾)骨上(圖 8-1)。甲附屬器始於遠端指(趾)骨皮膚表皮在稱為甲上皮 (eponychium) 的界線處的延伸(圖 8-2)。在該處,皮膚表皮向腹側內折並轉變為甲表皮 (nail epidermis),形成甲近端褶 (nail proximal fold, NPF)(見圖 8-2)。有趣的是,正常的表皮分化在近端褶於甲上皮界線處向內折之後便停止;因此,越過此點繼續延伸的甲層並不會形成正常皮膚表皮中通常可見的顆粒層 (granular layer)。作為近端褶在向背側與遠端彎折之後的腹側延續,便是甲基質 (nail matrix),由稱為甲母細胞 (onychocytes) 的活躍增生甲細胞所組成(見圖 8-2 與 8-3)。¹ 在甲基質上方為角化生成區 (keratogenous zone, KZ),基質細胞在此分化、扁平化、死亡,並沉積到其上方覆蓋的甲板中(見圖 8-2 與 8-3)。甲板 (nail plate) 是一個堅硬的結構,作為保護性覆蓋,防止腳趾與手指尖端受到外傷;它也可作為拾取小物體的工具,這對於精細操作與微妙的手指功能十分重要。透過甲板,可見白色新月形的基底稱為甲半月 (lunula,「小月亮」),是甲基質可見的部分(見圖 8-1)。甲半月在拇指上最為明顯,在小指上可能無法看見。
2
在甲器官內,甲板含有一個堅硬的角化結構,由稱為角質細胞 (corneocytes) 的扁平、無核細胞所組成,覆蓋於遠端指(趾)骨之上(見圖 8-1 與 8-2)。角質細胞是在甲分化過程中由甲細胞 (onychocytes) 形成的。甲板在指尖施加反向壓力以提供保護並增強敏感度。甲基質遠端為甲床 (nail bed),由基底層 (basal layer) 與一至兩層基底上方的有絲分裂後角質細胞 (suprabasal postmitotic keratinocytes) 所組成,貢獻少數角化細胞 (horn cells) 至遠端甲板的下表面(見圖 8-2)。² 甲板透過與其下方甲床交錯嵌合而附著於手指(見圖 8-2)。甲角皮 (nail cuticle) 由近端褶的邊緣延伸到甲板上,封閉指甲的近端,並保護其免受毒素與外來物質侵入(見圖 8-1)。在兩側,甲板被外側甲溝 (lateral nail grooves) 與外側甲褶 (lateral nail folds) 所環繞(見圖 8-1)。同樣地,在甲單位的遠端界線處,下甲皮 (hyponychium) 在甲皮帶 (onychodermal band) 下方將甲板封閉至甲床,以防止感染(見圖 8-1 與 8-2)。將人類指甲與其他哺乳動物的指甲(例如本例中的小鼠指甲)相比,可明顯看出形狀有很大不同,這很可能是由於演化與功能上的差異。人類指甲較為扁平(見圖 8-1 與 8-3);哺乳動物的爪則是一個近似圓錐形、曲度較大的結構。此形狀受到其下方遠端指(趾)骨以及甲基質的分布所影響。¹,³
甲生長與分化 (NAIL GROWTH AND DIFFERENTIATION)
到目前為止,一般認為甲基質主要負責甲板的生成。然而,先前並不清楚甲基質是否為甲分化的唯一來源,或甲單位的其他部分(例如甲床)是否也對甲板有所貢獻。過去,Lewis 觀察到甲板由三層不同的層次組成,並提出它們源自甲單位的三個不同部分。當時的理論認為,背側甲的淺層源自甲近端褶 (proximal nail fold),中間甲源自甲基質,而深部腹側甲則由甲床所生成。⁴
約十年後,Zaias 與 Alvarez 使用放射性氚標記甘胺酸 (radioactive tritiated glycine) 來標記並追蹤松鼠猴的甲細胞,證明該標記的攝取會隨時間從甲基質移動至甲板。⁵ 甲床中僅併入少量的放射性氚標記甘胺酸,因此因其不活躍而被排除為甲生成的來源。綜合來看,甲基質被提出為唯一負責甲板形成者。⁵ 類似的研究也在人類志願者身上進行,同時使用氚標記胸腺嘧啶核苷 (³H-thymidine) 與甘胺酸,研究中提出甲基質細胞除了遷移至甲板外,也會遷移至甲床。⁶ 「指甲僅由甲基質生成」的假說,進一步獲得 Berker 與 Angus 的支持,他們證明甲基質細胞相較於相對不活躍的甲床具有高度增生性。⁷ 在其他研究中,有人提出雖然甲基質生成甲板的大部分,但根據甲厚度與質量,甲床也對甲板有所貢獻。⁸,⁹ 有論點認為甲床對腹側甲板的貢獻,使甲板在生長時能向遠端移動。因此,總結而言,目前似乎存在一個普遍共識,即甲板是由甲基質細胞所生成的。
甲器官中存在兩個幹細胞庫(恆定與再生)(TWO POOLS OF STEM CELLS EXIST IN NAIL ORGAN [HOMEOSTASIS AND REGENERATION])
本節評估的是:甲基質細胞是否為甲生成所需前驅細胞的唯一來源,或是包括 NPF 在內的其他層次是否也在一生中貢獻部分指甲,以及它們本身是否符合作為幹細胞的標準。在全身各種器官中發現的成體幹細胞 (adult stem cells),負責維持器官的正常更新以及在受傷時的組織修復。¹⁰ 這些功能得以實現,是因為它們具有自我更新並分化為其各自器官中不同特化細胞類型的能力。¹⁰
由於幹細胞在一生中都被組織恆定與再生所需要,它們必須具有長期自我更新的能力。此外,有人提出幹細胞分裂頻率低,以避免併入與細胞分裂相關的突變。在每個器官中,幹細胞都駐留在稱為微環境 (niches) 的特化微環境中,這有助於維持靜止狀態或調控增生與分化,這對於組織恆定與修復十分重要。¹⁰⁻¹³
活化後,這些幹細胞分裂並離開微環境,形成分化程度較高的過渡放大型 (transit-amplifying, TA) 前驅細胞。相較於相對靜止的幹細胞,TA 細胞會快速增生並分化為再生或修復所需的細胞。¹⁰ 成體幹細胞理論上存在於所有可再生的組織中,但由於對這些器官所知甚少,其定位與特性描繪往往十分困難。因此,科學家利用慢循環的特性來標記並辨識各種器官中假設性的幹細胞。在早期的脈衝追蹤 (pulse-chase) 研究中,動物被注射 ³H-thymidine,它會併入分裂細胞新合成的 DNA 中。在有效標記後,動物經歷一段追蹤期,此時分裂的細胞會稀釋掉 ³H-thymidine 標記,而慢循環細胞則保留此放射性標記。此方法使得辨識慢循環的標記保留細胞 (label-retaining cells, LRCs) 成為可能。1990 年,Cotsarelis 及其同事使用 ³H-thymidine 脈衝追蹤實驗,在毛囊 (hair follicle) 的隆突區 (bulge region) 辨識出一個獨特的慢循環 LRCs 族群。他們證明這些隆突 LRCs 在以腫瘤促進劑(12-O-tetradecanoylphorbol-13-acetate [TPA])處理後可被刺激增生,並進一步假設這些 LRCs 是毛囊的幹細胞。¹⁴ 在辨識出毛囊隆突中的 LRCs 之前,Cotsarelis 及其同事也使用類似的脈衝追蹤實驗報告了角膜緣 (cornea limbus) 中的 LRCs。¹⁵ 這些角膜緣 LRCs 同樣會在受傷時被優先活化,也可被 TPA 刺激。
這些數據支持「幹細胞在動物的一生中持續存在,以在再生與傷口修復期間貢獻組織」的理論。然而,使用放射性標記時,研究人員無法透過免疫組織學探查潛在標記分子的共定位。不久之後,科學家開始使用一種新的合成胸腺嘧啶核苷類似物——溴去氧尿苷 (bromodeoxyuridine, BrdU)。¹⁶ 與 ³H-thymidine 相反,BrdU 不具放射性。此外,抗 BrdU 抗體使免疫組織化學能夠探查與 BrdU 標記的 LRCs 共定位的標記分子。¹⁷ 使用 BrdU 的脈衝追蹤研究已被廣泛用於辨識眾多組織中的 LRCs。最近,有人描述了一種使用四環素誘導型組蛋白 2B-綠色螢光蛋白 (tetracycline-induced histone 2B-green fluorescent protein, H2B-GFP) 轉殖基因小鼠來辨識 LRCs 的新方法。¹⁸ 透過此系統,可使用特異性或普遍性啟動子來驅動目標組織中可誘導的 H2B-GFP 表現。此系統的一項優點是它能在追蹤前更有效地標記細胞,而 BrdU 方法只能標記在脈衝期間於合成期 (synthesis phase) 中主動分裂的細胞。此外,此 H2B-GFP 轉殖基因標記保留系統允許分離活的 LRCs 以進行進一步的特性描繪與分析。此方法首次能夠分離活的毛囊標記保留幹細胞,以進行基因表現分析,從而獨立於任何其他標記分子(如角蛋白 15 [keratin 15, K15] 或 CD34)來確定這些幹細胞的分子特徵。¹⁸ 多年來,此方法使人們得以了解毛囊及其幹細胞是如何被調控的。
甲器官中慢循環的雙功能甲近端褶幹細胞 (SLOW-CYCLING BIFUNCTIONAL NAIL PROXIMAL FOLD STEM CELLS IN THE NAIL ORGAN)
隨後,除了毛囊外,在許多皮膚附屬器中(如角膜與汗腺),也證實了慢循環幹細胞的特性 ¹⁴,¹⁵,¹⁹⁻²¹;然而,對於指甲等其他皮膚附屬器則所知甚少。因此,近期研究嘗試探討:在持續生長的指甲中是否也存在這樣一個靜止的幹細胞族群。在小鼠指甲中,經 BrdU 脈衝追蹤實驗後,首次注意到指甲中 LRCs 的存在,顯示 LRCs 存在於與甲床相鄰的甲基質基底層中;然而,未進行進一步的特性描繪。²² 與此定位相反,Sellheyer 及其同事使用免疫組織化學染色,顯示人類指甲的腹側近端褶 (ventral proximal fold) 在胚胎發生 (embryogenesis) 期間表現毛囊幹細胞 (hair follicle stem cell, hfSC) 標記分子(例如 K15;角蛋白 19 [keratin 19, K19];以及 pleckstrin 同源樣結構域家族 A 成員 1 蛋白 [pleckstrin homology-like domain, family A, member 1 protein, PHLDA1]),且含有極少數 Ki67⁺ 細胞(Ki67 是與細胞增生相關的核蛋白),使它們更趨於靜止,類似於 hfSCs,從而提示近端褶可能代表人類 NSC 的微環境。²³
2
這些不一致之處已在近期研究中得到解決,這些研究使用轉殖基因小鼠的 H2B-GFP LRCs 系統,以在體內偵測靜止、慢循環的皮膚細胞。²⁴ 此方法能在 NPF 的基底層內辨識出一個先前未曾報導過的甲 LRCs 族群(圖 8-4)。²⁴ 有趣的是,在小鼠中,指甲以三維近似圓錐形的結構存在;因此,LRCs 以環狀構型排列。²⁴ 甲 LRCs 表現 hfSC 標記分子 K15,且體內譜系追蹤 (lineage tracing) 實驗顯示,源自 NPF 的 K15 標記細胞,同時貢獻於甲結構(見圖 8-4,黑色箭頭 #2,長期)以及更主要地貢獻於甲周表皮 (perinail epidermis)(見圖 8-4,綠色箭頭 #1),因此具有雙功能幹細胞 (bifunctional stem cell) 的特性。在甲再生時,這些 K15 衍生的 NPFSCs 主動將子代細胞輸送至甲基質並分化為甲板(見圖 8-4,黑色箭頭 #2)。同樣地,體內移植 (engraftment) 實驗證明甲 LRCs 能主動參與功能性的甲再生。對分離的甲 LRCs 進行轉錄分析揭示,正確的甲形成與分化需要 BMP 訊息傳導(圖 8-5)。因此,BMP 訊息傳導引導 NPFSCs 朝向甲分化,若缺少此路徑,甲基質細胞增生減少,且在甲基質區域上方觀察不到 KZ(見圖 8-5)。²⁴ 此外,正確的甲板分化受到損害,缺少 BMP 訊息傳導時,指甲在體內採取表皮命運 (epidermal fate)(見圖 8-5)。它表現為皮膚表皮顆粒層延伸遍及整個指甲,並在甲板中觀察到典型的表皮標記分子,如角蛋白 1 (keratin 1, K1) 與 loricrin 的表現(見圖 8-5)。相反地,典型的甲板分化標記分子 AE13(一種在毛皮質 [hair cortex] 與甲板中皆有表現的角蛋白)則無法偵測到(見圖 8-5)。²⁴ 有趣的是,在正確的毛囊分化中也觀察到 BMP 訊息傳導的類似角色。²⁵ 此觀察結果也一致,且其表型甚至比 Msx2 與 Foxn1 雙重突變體 (double mutant) 的表型更為嚴重;先前已證明這兩個基因都是 BMP 訊息傳導的下游標靶,調控正常的甲分化。²⁶ 它也類似於 Foxn1 與 Hoxc13 單一突變體,於甲結構內出現表皮顆粒層 (stratum granulosum) 的異常延伸(見圖 8-5)。²,²⁷ 此外,BMP 訊息傳導的角色也是維持 NPFSCs 之 LRC 特性所必需的(見圖 8-5),如同先前在 hfSCs 中所觀察到的。²⁸ 綜合來看,NPF 區域內一個新的雙功能幹細胞族群展現出可塑的恆定動態,能夠對損傷做出反應,並提示在指甲與相鄰表皮之間可能存在一種共同、協調的保護性屏障形成機制。²⁴
109
2
甲器官中快循環的單能性甲幹細胞 (RAPIDLY CYCLING UNIPOTENT NAIL STEM CELLS IN THE NAIL ORGAN)
甲基質是 NPF 細胞在形態學上的延續,這些細胞透過 IZ 彼此附著,包繞著甲板的近端(見圖 8-4)。因此,增生中的甲基質前驅細胞位於 NPF 區域的鄰近處,並分化為 KZ 的細胞以形成外部的甲板。在細胞增生動態方面,僅追蹤 1 週後,便可觀察到 NPF 中慢循環的甲 LRCs 與活躍分裂的甲基質細胞之間有明顯區別。²⁴ 在該時間點,Ki67 免疫標記主要定位於增生中的甲基質細胞,以及緊鄰於 Ki67 陰性、強 H2B-GFP⁺ 的近端褶 (proximal fold, PF) LRCs 旁的少數弱 H2B-GFP⁺ 細胞,顯示相較於相鄰的 PF 區域,甲基質內的增生增加(見圖 8-4)。因此,兩個區域之間的區域——增生中的甲基質與靜止的 PF IZ——被描述為含有弱 H2B-GFP⁺ 與 Ki67⁺ 的細胞,這些細胞源自細胞分裂後 H2B-GFP 標記的稀釋。IZ 隨著追蹤期延長而逐漸縮小,在追蹤 2 週後達到最小,並在追蹤 4 週後完全消失。這顯示存在一個細胞分裂梯度,從 NPF 中相對靜止的慢循環 LRCs(經過較活躍細胞的 IZ)擴展到快速分裂的甲基質細胞(見圖 8-4)。²⁴
因此,已證明甲基質含有負責甲分化的快速分裂前驅細胞。然而,甲基質細胞本身是否擁有幹細胞特性並能生成整個甲附屬器,這一點已被研究,並將在本章稍後進一步探討。為了定位指甲中的幹細胞,Takeo 及其同事使用了一個轉殖基因小鼠譜系追蹤系統。在角蛋白 14 (keratin 14, K14) 啟動子驅動的 Cre 重組酶-突變雌激素受體 (Cre recombinase–mutated estrogen receptor, Cre-ER)(K14–Cre-ER)的控制下,他們透過活化 LacZ 表現來標記皮膚表皮與甲表皮基底層中的角質細胞。²⁹ 在該系統中,LacZ 表現由 Rosa26 啟動子驅動,其活化發生在 Cre 介導的 floxed 終止匣 (stop cassette) 被移除之後。因此,它能以單次注射 tamoxifen 來基因性地標記一小群 K14 甲基底表皮細胞(包括甲基質細胞與甲床細胞),然後在一段較長時間內評估 LacZ 標記細胞在甲再生中的參與情形。在超過 5 個月的時間裡,被標記的 K14 甲上皮細胞的 LacZ⁺ 後代以線性方式向遠端延伸,以條紋形式持續存在,且這些條紋僅源自近端甲基質而非遠端甲基質。近端甲基質細胞具有未分化表皮細胞的特性,除了 K14 外還表現角蛋白 17 (keratin 17, K17),並具有高度增生的 Ki67 標記(見圖 8-4)。分離出的近端甲基質細胞在體外具有最高的群落形成能力 (colony-forming ability),這是上皮幹細胞的一般特性。因此,長期標記結果連同體外培養數據證明,快速分裂的近端甲基質含有自我更新的細胞,符合本身即為維持甲生長之 NSCs 的標準(見圖 8-4)。²⁹
有趣的是,近端甲基質與遠端甲基質之間的基因表現分析揭示,近端甲基質細胞富含 Wnt 訊息傳導路徑下調的 NSCs。²⁹ 事實上,使用兩種不同 Wnt 報告基因小鼠的分析證實,雖然 Wnt 訊號從 K17 陽性 NSC 區域的遠端部分開始,並持續進入 K17 陰性遠端甲基質的遠端基質中,但這兩種訊號在甲基質的近端皆不存在。一致地,Wnt 訊息傳導路徑的組成成分,如 Tcf1(亦稱為肝細胞核因子 1a [hepatocyte nuclear factor 1a],一種 Wnt 訊息傳導的核介導因子)與 Wls(Wntless 同源物 [Wntless homologue]),在 NSCs 的近端並不表現,但數個含有 TCF1 與 LEF1 共識結合位點 (consensus binding site) 的角蛋白則在遠端甲基質中上調,因此提示 Wnt 訊息傳導直接參與甲分化。這一點透過在甲上皮中刪除 Wnt 訊息傳導活化(在成體上皮中移除 β-catenin,一種 Wnt 訊息傳導的必要介導因子)而得到驗證,表現為缺乏 AE13(一種角化甲細胞的標記分子)。有趣的是,缺少 β-catenin 或 Wntless 時,整個甲上皮都表現出 NSC 區域的特性,即 K17 陽性與高度增生的 Ki67 陽性細胞,從而確認了 Wnt 訊息傳導在甲分化中的必要角色。此外,在近期一項研究中,科學家辨識出一個表現 Wnt 訊息傳導介導因子 Lgr6(富含白胺酸重複序列的 G 蛋白偶聯受體 6 [leucine-rich repeat containing G protein–coupled receptor 6])的細胞族群,作為指甲的關鍵幹細胞標記分子,其定位於近端甲基質內,因此與 Takeo 及其同事先前的發現一致並加以確認。³⁰ 事實上,Lgr6 是 NSCs 特有的分子標記,產生甲板並隨時間貢獻於指甲的生長(見圖 8-4)。因此,這些數據確認了典型 Wnt 訊息傳導(其導致 β-catenin 定位於甲基質細胞核中)與表現 Lgr6 的甲基質細胞的表現大致相關。
甲上皮在指(趾)端再生中的角色 (ROLE OF NAIL EPITHELIUM IN DIGIT REGENERATION)
人類指(趾)端,與包括小鼠在內的一些哺乳動物相似,能在受傷後像兩棲類一樣完全再生。³¹,³² 有趣的是,此能力僅限於與指甲相關的區域,先前研究顯示,在中節指(趾)骨 (middle phalanx) 截肢後進行甲移植可誘導異位的指(趾)骨分化。³¹⁻³³ 因此,此觀察結果提示甲上皮具有一項特殊功能,為指(趾)端再生創造一個許可性的再生環境。最近,Takeo 及其夥伴發現了指(趾)端再生過程的一些細胞與分子組成。²⁹ 事實上,此過程是經過協調的,並取決於上方覆蓋的甲器官的存在及其與下方胚芽 (blastema) 的相互交互作用。已證明甲上皮,特別是位於近端甲基質的 NSCs 及其分化,與協調指(趾)端再生的能力直接耦合。在分子層次上,於小鼠模型中,已證明來自甲上皮的 Wnt/β-catenin 訊息傳導是指(趾)端再生所必需的。目前提出的模型是:近端甲基質的 NSCs 透過 Wnt 活化(經由 Wntless)產生遠端甲基質細胞,同時,NSCs 與遠端甲基質細胞分化為甲板(圖 8-6)。在表現 Wntless 的遠端甲基質層級進行截肢(但近端甲基質的 NSCs 保持完整)後,傷口部位被再生中的甲上皮細胞所覆蓋,這些細胞接著活化 Wnt 訊息傳導(Wntless)以分化為遠端甲基質細胞與甲板(見圖 8-6)。Wnt 路徑的活化透過 semaphorin 5a(Sema5a,一種軸突導引分子 [axon-guidance molecule])促進胚芽神經支配 (blastema innervations),Sema5a 在截肢後 3 週的對照組甲上皮中上調。反之,神經支配是再生中甲上皮表現纖維母細胞生長因子 2 (fibroblast growth factor 2, FGF2) 所必需的(見圖 8-6)。接著,FGF2 促進 Runx2 陽性前驅細胞或 Sp7 成骨細胞 (osteoblasts) 的增生,最終導致耦合的指(趾)端再生(見圖 8-6)。有趣的是,FGF2 配體的受體 FGFR1 表現於間質胚芽 (mesenchymal blastema)(一種 Runx2 前驅細胞)中;因此,ERK 路徑可能被招募以活化並維持其增生(見圖 8-6)。因此,甲分化由一個 WNT 依賴性機制所調控,此機制與指(趾)端再生相連結。事實上,Wnt 刪除(在 β-catenin 條件性基因剔除 [conditional knockout, KO] 小鼠中)導致甲分化缺失,表現為甲基質中缺乏 TopGal 表現(一種 WNT 路徑的基因報告子)與 AE13 表現(一種角化甲板細胞的標記分子),且骨再生受阻。此外,在 β-catenin 條件性 KO 小鼠中,甲基質上皮下方的 Runx2 前驅細胞、間質細胞與 Sp7 成骨細胞並未被刺激增生或產生 BMP4。再者,Wnt 依賴性的神經支配可促進指(趾)端再生,因為在 β-catenin 條件性 KO 小鼠中,截肢後 3 週甲上皮中的 Sema5a 下調。這些數據一致顯示,去神經支配 (denervation)(在截肢前以手術移除神經)會抑制胚芽生長,類似於在缺乏 FGF2 表現的甲上皮去神經支配後於條件性 KO 小鼠中所觀察到的情形。³⁴,³⁵ 有趣的是,雖然在近端截肢甲基質細胞後,β-catenin 的穩定化會誘導 TCF1 表現以及遠端甲基質的再生和良好神經支配的胚芽細胞之形成,但在 NSC 微環境近端進行截肢後,甲上皮中 β-catenin 的穩定化並不促進指(趾)端再生。因此,剩餘的近端甲基質表皮無法對 β-catenin 穩定化做出反應,指(趾)端再生無法恢復,這與遠端甲基質的功能形成對比。此結果與消融 Lgr6(Wnt 路徑的一個重要促效劑,標記 NSCs 並產生甲板)後正確的指(趾)端再生受抑制相一致。³⁰ 此外,Lgr6 表現不僅限於甲基質,更廣泛地表現於指(趾)端的一群細胞中,亦即骨成骨細胞 (bone osteoblasts) 與外分泌汗腺 (eccrine sweat glands) 中。因此,表現 Lgr6 的細胞不僅標記 NSC 上皮,也貢獻於胚芽,這提示表現 Lgr6 的細胞在指(趾)端再生過程中不僅有直接角色,也有潛在的間接角色。³⁰ 事實上,此角色已透過對 Lgr6 缺陷小鼠的分析得到證實,這些小鼠同時有指甲與骨再生的缺陷,但 Lgr6 陽性成骨細胞是否貢獻於此表型仍不清楚。因此,這些細胞在正常骨骼恆定與指(趾)端再生中的角色仍有待未來確定。總體而言,Wnt 訊息傳導在 NSC 譜系中的雙重功能——指揮甲形成與分化以及指(趾)端再生——似乎是協調哺乳動物指(趾)端再生中上皮與間質組織再生的關鍵機制(見圖 8-6)。事實上,有趣的是,一些人類中可能僅影響指甲的甲疾病包括遺傳性無甲症 (inherited anonychia) 與孤立性先天性甲發育不良 (isolated congenital nail dysplasia)。罹患遺傳性無甲症的家族有嚴重的甲發育不全 (hypoplasia),其中辨識出 R-spondin 4 (Rspo4) 的突變,Rspo4 與 Wnt 訊息傳導路徑有關,提示 Rspo4 在甲發育中的重要角色。³⁶
結論 (CONCLUSIONS)
幹細胞具有長期自我更新的潛能,並能分化為多種不同的細胞類型。這種持續性與多能性對於在恆定與修復期間維持組織至關重要。由於其非凡的再生潛能,了解其維持與調控將證明其在組織再生與各種疾病治療中的有用性。目前的發現揭示了一個模型,在該模型中,甲器官中的幹細胞以兩種不同的活化模式存在——慢循環與活躍增生——以滿足持續生長的指甲之需求。²⁴,²⁹ 這些數據證明甲器官含有一個由慢循環到快循環幹細胞的梯度,包括近端褶中慢循環的 NPFSCs、IZ 中較活躍的細胞,以及最終近端甲基質區域中快速增生的 NCSs。總體而言,此模型支持目前關於成體幹細胞在多個器官與組織中於恆定與再生期間之動態與能力的觀點。例如,標記保留的 NPFSCs 與快速增生的 NCSs 呈現出一種類似於 hfSCs 中所描述的動態階層,在 hfSCs 中,一些 hfSCs 參與新的毛髮週期,而生長毛囊上部外根鞘 (upper outer root sheath, ORS) 的其他 hfSCs 則以靜止的 hfSCs 重建新的隆突。³⁷
在此隆突活化期間,可在 ORS 內觀察到一個由慢循環到快循環細胞的梯度,分別從隆突區延伸向甲基質。為何甲器官擁有兩個幹細胞族群?一個答案可能來自於對腸道中具有不同細胞週期動態的兩個幹細胞族群之類似系統的觀察。在腸道中,慢循環的 LRCs 幹細胞已被辨識位於腸隱窩 (intestinal crypts) 的 +4 位置,表現 Bmi1。³⁸,³⁹ 相反地,快循環的幹細胞也在隱窩底部被發現,表現 Lgr5,⁴⁰ 且這兩個幹細胞族群都能分化為所有腸道譜系。³⁹ 有趣的是,最近發現,在缺乏快速增生的 Lgr5⁺ 幹細胞時,Bmi1 幹細胞能維持腸道的正常更新,同時重新填補 Lgr5⁺ 幹細胞族群。⁴¹ 這提示位於 LRCs 所在位置的 Bmi1⁺ 細胞作為一個儲備幹細胞族群。因此,指甲中兩個幹細胞族群(即 NPFSCs 與 NSCs)之間可能存在一個類似的備份系統,以保護整個甲器官的維持。因此,未來進一步探討這一點將會很有意思。總體而言,這提示近端甲基質中的 NSCs²⁹ 整合了相鄰的近端褶作為雙功能 NPFSCs 的所在位置。²⁴ 綜合來看,這些範例突顯了甲上皮內成體幹細胞族群的內在可塑性,而這些高度增生組織中慢循環的 NPFSCs 可能代表 NSCs 在壓力、損傷或幹細胞族群耗竭期間的備份系統。所有這些關於甲器官中幹細胞族群及其相互關係,以及幹細胞與幹細胞微環境之間相互交互作用的令人振奮的發現,有助於揭示幹細胞生物學的新概念,並具有可能大幅影響再生醫學的偉大轉譯潛能。事實上,在小鼠指(趾)端再生中已證明,雖然帶有幹細胞的甲上皮對於協調此過程至關重要,但與其他胚層及譜系受限的幹/前驅細胞之間的交互對話,對於成功完成此再生是必不可少的。⁴² 因此,未來這項研究可能有潛力透過協調完整指(趾)端再生(包括骨、神經支配與血管,連同皮膚與皮膚附屬器以及指甲)來恢復解剖學與功能性的結構,從而造福截肢的個體。這些結果已建立了 NSC 分化與指(趾)端再生之間的直接關係,並提示 NSCs 可能具有治療潛能,能貢獻於開發針對截肢者的新型治療。這是一個非常重要的問題,因為全世界有許多人受到手指或腳趾意外截肢的影響。近期的甲與指(趾)端再生數據已開始為揭示開發此再生潛能所需的潛在機制鋪路。因此,甲生物學的新發現很可能會增進我們對協調指(趾)端再生所需基本機制的理解,並有潛力解鎖超越指(趾)端再生、重建整個肢體的廣泛再生潛能。雖然仍需更多研究以進一步描繪甲器官中這些幹細胞族群的特性,但這些數據可能在未來為人類指甲假設性的幹細胞標記分子帶來一些啟發。事實上,有趣的是,在人類中最初的觀察提出,腹側近端褶基於表現隆突幹細胞標記分子 K15、K19 與 PHLDA1 並具有靜止特性,是發育中的胚胎期人類指甲的 NSCs,這可能對應於小鼠模型中的 NPFSCs。²³ 相反地,人類甲基質含有以 Ki67 標記的高度增生細胞,類似於小鼠甲基質,這可能類似於小鼠近端甲基質中高度增生的 NSCs 幹細胞。²³ 因此,進一步辨識、描繪特性並分離人類甲器官中腹側近端褶與甲基質這兩個不同的幹細胞族群,可能會在分子層次上提供對甲器官生物學的關鍵理解,這有潛力為未來設計重新生長更大部分肢體以及其他不可再生組織的轉譯療法提供洞見。
113
2
因此,定義這些 NSCs 與 NPFSCs 的再生潛能,以及它們在體內被調控的分子機制(期望藉此出現新型療法),可能會朝向成功的甲器官再生與傷口癒合(包括截肢病人的治療)發展。
致謝 (ACKNOWLEDGMENTS)
我向那些其研究無法納入本章的同事們致上歉意。我感謝 Sébastien de Feraudy 醫師(MD, PhD,UC Irvine 皮膚病理學實驗室副教授兼主任,加州 Irvine),他提供了人類指甲切片後的蘇木精-伊紅 (hematoxylin and eosin) 染色圖。我感謝 Agnieszka Kobielak 博士對本手稿的批判性審閱,以及 Krzysztof Kobielak 實驗室成員的有益討論。KK 受到國家科學中心 (National Science Centre, NCN) Opus Grant 2015/19/B/NZ3/02948 以及美國國家衛生研究院之國家關節炎與肌肉骨骼及皮膚疾病研究所 (National Institute of Arthritis and Musculoskeletal and Skin Diseases of the National Institutes of Health) Grant R01-AR061552 的支持。
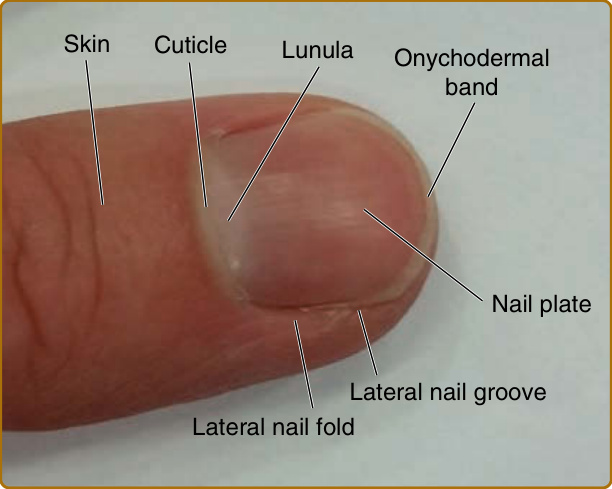
圖 8-1:人類指尖與甲器官的圖片。可見的結構包括皮膚、角皮 (cuticle)、帶有近端甲半月 (lunula) 的甲板,以及位於遠端的甲皮帶 (onychodermal band)。在側邊,甲板被外側甲溝 (lateral nail groove) 與外側甲褶 (lateral nail fold) 所分隔。
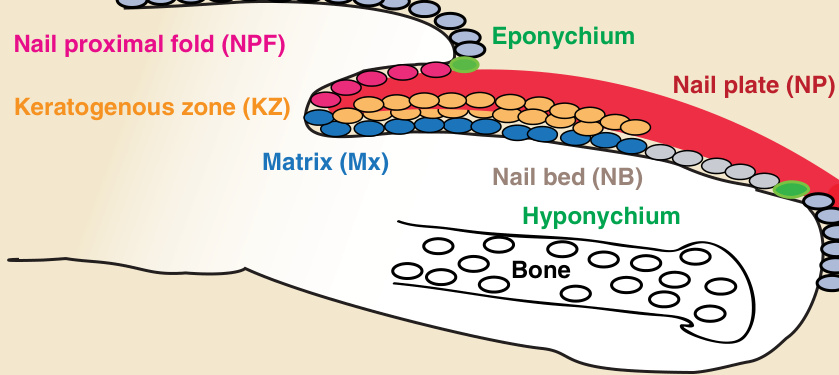
圖 8-2:人類指甲結構示意圖(通過指甲中央的縱切面)。以顏色標示的不同甲層與結構包括:甲近端褶 (nail proximal fold,粉紅色)、甲基質 (matrix,藍色)、角化生成區 (keratogenous zone,橘色)、甲床 (nail bed,淺灰色)、甲板 (nail plate,紅色),兩側由甲上皮 (eponychium) 與下甲皮 (hyponychium,綠色) 包夾,後者將甲結構與皮膚表皮 (skin epidermis,深灰色) 分隔開來。
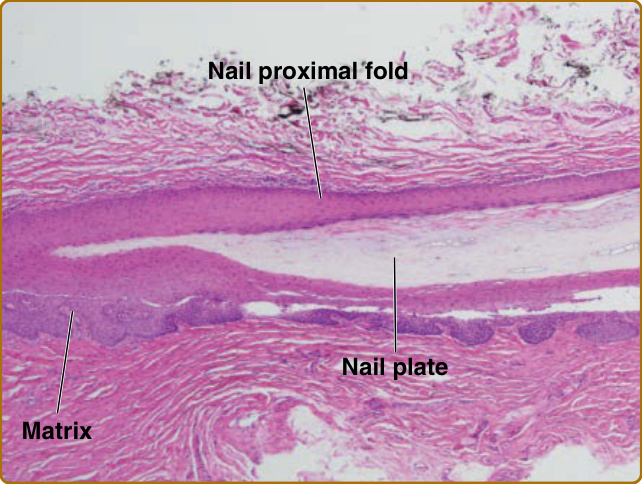
圖 8-3:甲器 (nail apparatus)。構成甲單位之各元素的組織學外觀(通過指甲中央縱切面的蘇木精-伊紅 [hematoxylin and eosin] 染色;原始放大倍率 ×20)。甲的組織學層次為甲近端褶、甲基質與甲板。(經 Sébastien de Feraudy 醫師許可使用,MD, PhD,UC Irvine 皮膚病理學實驗室副教授兼主任,加州 Irvine。)
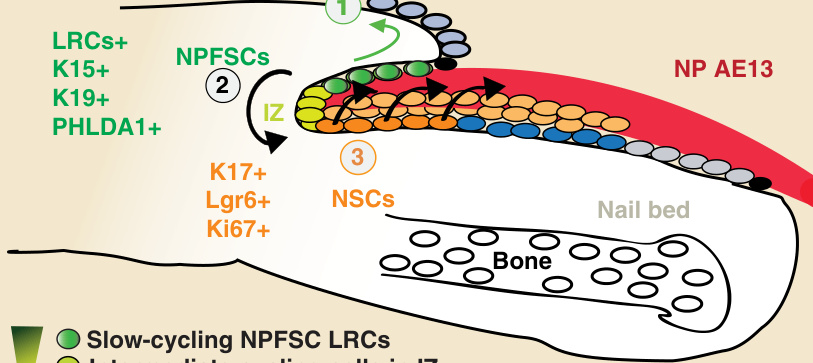
圖 8-4:指甲中兩個幹細胞族群的定位示意圖。具有慢循環標記保留特性 (label-retaining characteristic, LRCs) 的甲近端褶幹細胞 (nail proximal fold stem cells, NPFSCs) 定位於甲近端褶,而快速增生的甲幹細胞 (nail stem cells, NSCs) 定位於近端甲基質。NPFSCs 表現角蛋白 15 (keratin 15, K15);角蛋白 19 (keratin 19, K19);以及 pleckstrin 同源樣結構域家族 A 成員 1 蛋白 (PHLDA1)。K15 標記的 NPFSCs 在甲恆定期間主要貢獻於甲周表皮 (perinail epidermis)(1,綠色箭頭),但也貢獻於甲基質與甲分化(2,黑色箭頭)。NSCs 當以 Lgr6 標記時,參與甲基質結構與甲板 (nail plate, NP) 分化(3,黑色箭頭)。AE13,於毛皮質與甲板中表現的角蛋白;IZ,中間區 (intermediate zone);Ki67,與細胞增生相關的核蛋白;Lgr6,富含白胺酸重複序列的 G 蛋白偶聯受體 6。
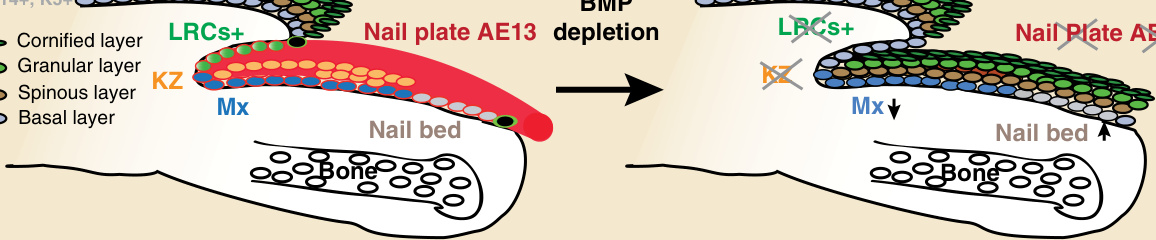
圖 8-5:骨形態發生蛋白 (bone morphogenetic protein, BMP) 訊息傳導是正確甲分化所必需的。活躍的 BMP 訊息傳導見於甲近端褶與甲基質(以紅色框圈標示)。指甲中的 BMP 耗竭證明角化生成區 (keratogenous zone, KZ,橘色圈) 缺失、甲床增生 (hyperplasia,灰色圈)、標記保留細胞 (label-retaining cells, LRCs) 喪失,且異位的顆粒層延伸覆蓋整個甲板(以綠色圈標示的皮膚表皮層)。角蛋白 1 (keratin 1, K1) 與 Loricrin (Lor) 表皮標記分子在缺乏 BMP 訊息傳導時異位表現。在對照組小鼠中,陽性 K1 染色見於近端褶與腹側甲。觀察到異位的 K1 表現(以棕色圈標示的皮膚表皮層)取代了甲板標記分子 AE13(紅色)的表現。在對照組小鼠中,Loricrin 表現見於近端褶直至甲上皮,並在背側甲板中缺失。觀察到異位的 Loricrin 表現(以深綠色圈標示的皮膚表皮層)取代了甲板標記分子 AE13(紅色)的表現。Mx 中增生細胞的數量(以藍色圈標示)似乎減少,且缺乏 BMP 訊息傳導時,甲板分化的標記分子 AE13 受到損害。因此,表皮標記分子的異位表現證明 BMP 訊息傳導可能是甲命運所必需的,缺少它時,甲結構會獲得類似表皮的表型。
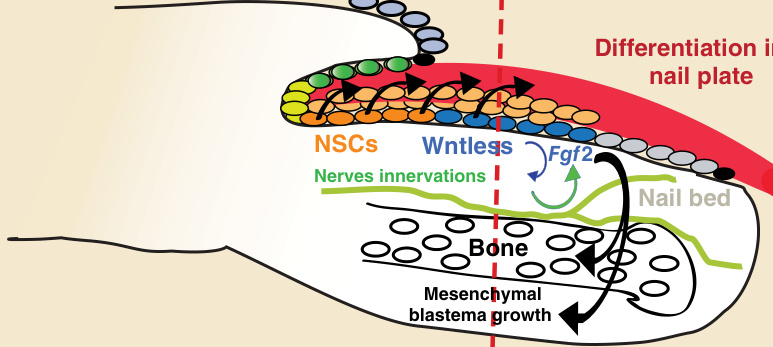
圖 8-6:甲上皮分化與指(趾)端再生之間的分子連結。近端甲基質中的甲幹細胞 (nail stem cells, NSCs,深橘色圈) 在恆定狀態下透過 Wnt 活化 (Wntless) 產生遠端甲基質細胞(藍色圈)。近端與遠端甲基質細胞皆分化為甲板(紅色)。在遠端層級截肢(紅色虛線)後,甲上皮細胞開始再生傷口部位,並伴隨 Wnt 訊息傳導的活化以分化為遠端甲基質細胞與甲板。甲上皮中此 Wntless 活化是促進(藍色箭頭)胚芽神經支配(blastema innervations,綠色)所必需的,而胚芽神經支配又是再生中甲上皮(藍色圈)表現 Fgf2 所必需的(綠色箭頭)。隨後,纖維母細胞生長因子 2 (fibroblast growth factor 2, FGF2) 促進(黑色箭頭)Runx2 陽性間質細胞的增生,最終導致指(趾)端再生。(改編自 Takeo M, Chou WC, Sun Q, et al. Wnt activation in nail epithelium couples nail growth to digit regeneration. Nature. 2013;499(7457):228-232.)