皮膚血管系統 (Cutaneous Vasculature)
PART 2
皮膚的結構與功能 (Structure and Function of Skin)
重點一覽 (AT-A-GLANCE)
■ 皮膚血管系統分為淺層 (superficial)、深層 (deep) 與皮下 (subcutaneous) 三個血管叢 (vascular plexus)。淺層血管叢由成對平行的小動脈 (arterioles) 與小靜脈 (venules) 構成,兩者透過延伸進入真皮乳頭 (dermal papillae) 的微血管環 (capillary loops) 相連。這些節段可單獨或共同對外源性或內源性刺激產生反應,從而影響皮膚疾病的表現。
■ 皮膚微血管由內皮細胞 (endothelial cell) 內襯構成,並由壁細胞 (mural cells) 支持。在大部分微血管系統中,壁細胞為周細胞 (pericytes),但在較大的小動脈與小靜脈中則為平滑肌細胞 (smooth muscle cells),並有駐留的血管周圍白血球 (resident perivascular leukocytes),包括 T 細胞、巨噬細胞 (macrophages)、肥大細胞 (mast cells) 與樹突狀細胞 (dendritic cells)。
■ 皮膚微血管與其他微血管一樣,執行三項重要的固有功能:調節血液的流動性 (fluidity)、形成一道分隔並控制分子與細胞在循環血液與組織之間轉移的屏障,以及調節局部血流。
■ 控制血流通過皮膚微血管在體溫調節 (thermoregulation) 中具有特殊且關鍵的角色,這是血管系統其他節段所不執行的功能。
■ 皮膚微血管細胞的型態、基因表現與功能會因急性或慢性發炎性皮膚疾病及癌症而改變。這些過程可能涉及新血管的形成(血管新生 angiogenesis)或既有血管的重塑 (remodeling)。
■ 皮膚特異性的微血管反應可能受到角質細胞衍生 (keratinocyte-derived) 及其他環境衍生因子的影響。
前言 (INTRODUCTION)
血液血管系統 (blood vascular system) 是一系列連續的中空管道,內襯一層僅一個細胞厚、類似上皮的間葉細胞——即內皮 (endothelium)——並由各種壁細胞支持,典型者為微血管中的周細胞 (pericytes, PCs) 與較大管徑血管中的平滑肌細胞 (smooth muscle cells, SMCs)。所有血管內皮細胞 (endothelial cells, ECs) 都具有共同的特徵與功能,因此可被集體描述為一種細胞類型。然而,血管系統某一節段的 ECs 在重要層面上可能與其他解剖部位的 ECs 有所不同。血管內皮細胞也不同於淋巴管內皮細胞 (lymphatic ECs),後者不在本章討論範圍內。壁細胞在整個血液血管系統中具有不同的胚胎學起源,但對於它們隨解剖位置而產生的變異所知甚少。因此,這些細胞將被視為同質的來討論,但對於此類描述應抱持審慎態度。
皮膚血管系統的結構與組織 (STRUCTURE AND ORGANIZATION OF THE SKIN VASCULATURE)
大約三十年前,Irwin Braverman¹ 描述了人類皮膚血管網絡的組織結構。人類真皮血管系統由兩個相互連接的系統所組成——淺層血管叢與深層血管叢 (deep vascular plexus, DVP),並有環繞汗腺 (sweat glands) 與毛囊 (hair follicles) 的額外血管網絡(圖 9-1)。淺層血管叢 (superficial vascular plexus, SVP) 由成對的小動脈與小靜脈構成,它們形成一個相互連接的血管網絡,在一個平行於並緊鄰表皮表面下方的平面上行進。微血管 (capillaries) 起源於小動脈,在表皮腳脊 (epidermal rete ridges) 之間的部位向上延伸進入乳頭層真皮 (papillary dermis),然後再向下迴繞回到小靜脈,形成拱廊狀 (arcade-like) 結構。在電子顯微鏡下,小動脈的基底膜 (basement membrane) 呈現均質外觀,而小靜脈的基底膜則為多層狀。在正常皮膚中,大部分的微血管環被一層類似小動脈的基底膜所包覆,僅在最深處腳脊、緊鄰與 SVP 小靜脈吻合處的近端,才取得類似小靜脈的包覆構造。皮膚不同部位的微血管環之間並無超微結構上的差異。SVP 的小動脈與小靜脈透過短而直的血管,連接至一個更深的平面狀吻合血管網絡的小動脈與小靜脈,此網絡稱為 DVP(見圖 9-1)。DVP 的平面平行於 SVP 的平面,並在網狀真皮 (reticular dermis) 與其下方皮下組織 (subcutis) 之間的交界處上方行進。DVP 的小動脈由來自皮下組織的穿通血管 (penetrating vessels) 供應。DVP 的小靜脈由含瓣膜的靜脈引流,這些靜脈回流至皮下脂肪。連接 DVP 小動脈與小靜脈的微血管網絡為網狀真皮內的附屬器構造提供營養。DVP 的小靜脈作為與附屬器發炎相關、或與網狀真皮本身發炎相關之白血球的進入門戶 (portals of entry)。真皮小動脈的管壁由三層構成:由 ECs 組成的內膜 (intima)、由 SMCs 構成的中膜 (media),以及含有一些結締組織型細胞的外膜 (adventitia)。在終末小動脈 (terminal arterioles) 中,SMCs 可能被位於 ECs 基底膜內(而非位於獨立中膜腔室內)的 PCs 所取代。靜脈與小靜脈的管壁具有相似的結構,但與對應的動脈與小動脈相比,靜脈循環的血管具有較大的管腔 (lumen)、較薄的肌肉壁,並在小血管連接至較大血管的部位設有瓣膜。真皮微血管與微血管後小靜脈 (postcapillary venules) 的管壁由一層位於簡單基底膜上的有窗孔 ECs (fenestrated ECs) 單層所構成(圖 9-2)。微血管典型上由單列高度彎曲的 ECs 內襯,這些 ECs 包圍管腔並與兩端相鄰的 ECs 緊密相連,形成一個管腔直徑通常小於 10 μm 的管道。微血管後小靜脈的管徑稍大,其環周由一個以上的 EC 內襯,相鄰 ECs 之間的連接較鬆散,並具有多層基底膜。真皮微血管 ECs 的窗孔 (fenestrae) 主要由一種稱為漿膜囊泡蛋白 (plasmalemmal vesicle protein, PV)-1 的蛋白質所形成,該蛋白先前被命名為 Pathologische Anatomie Leiden-endothelial (PAL-E) 抗原,並被用作微血管 ECs 的標記。位於真皮微血管基底膜內的 PCs 為微血管與微血管後小靜脈的管壁提供結構完整性,並參與基底膜的合成,以及(當其發生時)重塑。在皮膚微血管中,ECs 的數量多於 PCs,每個 PC 接觸多個 ECs 並形成黏著連接 (adherens junctions)。
SVP 的微血管後小靜脈也與各種駐留的白血球細胞群相關聯,包括 T 細胞、巨噬細胞、樹突狀細胞與肥大細胞。這些細胞連同 ECs 與 PCs,構成微血管後小靜脈的「血管周圍滲出單位 (perivascular extravasation unit)」²,這是血管系統中循環白血球可被招募進入真皮的節段。這些血管周圍駐留白血球群在二十多年前即已被辨識,而隨著對白血球譜系與發育程序的較新理解逐漸闡明,它們大多尚未被重新分類。例如,現今已認知記憶 T 細胞 (memory T cells) 可長期駐留於組織中,並獲得有別於循環 T 細胞群的特性與功能。³ 帶著這些新資訊,未發炎皮膚中的血管周圍 T 細胞群,尚未與循環 T 細胞或表皮駐留 T 細胞進行比較分類。樹突狀細胞現今已知具有相當的異質性,並擁有不同的發育程序;每種樹突狀細胞類型執行不同的免疫功能。同樣地,血管周圍樹突狀細胞的類型尚未依這些新標準加以定義。肥大細胞在其顆粒內容物與生物合成特徵方面也具有異質性,而人類真皮血管周圍肥大細胞可能合成什麼物質仍不清楚。駐留組織巨噬細胞 (resident tissue macrophages) 現今已知起源於較原始的胚胎來源(例如卵黃囊 yolk sac 或胎兒肝臟 fetal liver),或來自骨髓衍生的單核球 (bone marrow-derived monocytes),且這些細胞群也具有不同的特性。⁴ 最後,真皮樹突細胞 (dermal dendrocytes),又稱為帷幕細胞 (veil cells),形成一個駐留細胞群,包繞 SVP 的微血管,與 T 細胞及肥大細胞相關聯,並除了其他類巨噬細胞標記外,還表現凝血因子 XIIIa (coagulation factor XIIIa) 與穩定蛋白 1 (stabilin 1)。關於這些細胞究竟是骨髓衍生還是間葉衍生,一直存在許多爭論,而隨著對駐留巨噬細胞異質性的新認識,它們事實上可能兩者皆非。一般而言,有人提出所有此類血管周圍白血球細胞類型都作為感染或損傷的哨兵 (sentinels),但其中某些細胞群也可能具有固有的血管功能。真皮靜脈形成三種吻合連接:上行靜脈主幹的大型吻合 (large anastomoses)、小靜脈上行段中的小型吻合 (small anastomoses),以及從小型上行靜脈延伸至乳頭層真皮的極小型吻合 (very small anastomoses)。這個真皮靜脈的吻合網絡被組織成各種大小的多邊形 (polygons),被認為在皮瓣手術 (skin flap surgery) 的結果中扮演角色。
正常皮膚血管系統的特性與功能 (PROPERTIES OF AND FUNCTIONS OF NORMAL SKIN VASCULATURE)⁵
見表 9-1。
止血 (HEMOSTASIS)
血液是一種液體,在血管完整性受到破壞的部位準備轉變為凝膠 (gel),有益地形成血塊以限制外滲 (extravasation),但通常避免在血管管腔內形成可能阻塞血管並限制灌流 (perfusion) 的血栓 (thrombus)。血管 ECs 是負責維持血液流動性的主要細胞,並可在需要時被改變以主動促進血塊形成。血漿 (blood plasma) 含有凝血系統 (coagulation system) 的蛋白質,這些蛋白質產生纖維蛋白凝膠 (fibrin gels),以及可形成初級止血栓 (primary hemostatic plug) 的血小板 (platelets)。EC 內襯防止凝血系統與血小板的血管內活化,卻使這兩個系統都保持待命狀態以對損傷做出反應。ECs 基礎性表現的主要凝血抑制因子為:組織因子途徑抑制因子 (tissue factor pathway inhibitor)(可防止由組織因子 tissue factor 所催化、因子 VIIa 對因子 IX 與 X 之酵素活性的劇烈增加)、血栓調節蛋白 (thrombomodulin)(將凝血酶 thrombin 從切割並轉化纖維蛋白原 fibrinogen 為纖維蛋白 fibrin,重新導向為改而標靶並活化蛋白質 C protein C),以及抗凝血肝素硫酸鹽 (anticoagulant heparan sulfates)(活化抗凝血酶 III antithrombin III,使其作為凝血酶與因子 Xa 的抑制因子發揮功能)。ECs 也以下列方式基礎性地限制血小板活化:(1) 產生一氧化氮 (nitric oxide) 與前列環素 (prostacyclin,又稱前列腺素 I2 prostaglandin I2),透過分別提升細胞內環狀單磷酸鳥苷 (cyclic guanosine monophosphate) 與環狀單磷酸腺苷 (cyclic adenosine monophosphate) 來抑制血小板活化;(2) 表現可將活化血小板的三磷酸腺苷 (adenosine triphosphate) 與二磷酸腺苷 (adenosine diphosphate) 降解為惰性單磷酸腺苷 (adenosine monophosphate) 的外酶 (ectoenzymes);(3) 將凝血酶(透過切割蛋白酶活化受體 1 protease activated receptor 1 來活化血小板)的活化降至最低;以及 (4) 遮蔽基底膜與間質膠原蛋白 (interstitial collagens)(可被血小板表面蛋白辨識並作為血小板的替代活化劑)(圖 9-3)。
選擇性通透性 (PERMSELECTIVITY)
真皮 ECs 形成一道屏障,允許液體與溶質的有限通過,使血液能滋養組織,同時對巨分子 (macromolecules) 展現「選擇性通透性 (permselectivity)」(圖 9-4)。ECs 也基礎性地對血球細胞呈現一道幾乎無法穿透的屏障(除了在高通透性與白血球轉運的特化部位)。一般而言,小動脈與微血管對巨分子的通透性較低,因為內襯這些血管的 ECs 彼此之間透過緊密連接 (tight junctions) 相連,這些緊密連接含有閉合蛋白 (claudins,主要為 claudin 5)、連接黏附分子 (junctional adhesion molecules,主要為 JAM-A) 與閉合素 (occludin)(不過根據小鼠的基因破壞實驗,閉合素似乎是可有可無的)。緊密連接防止巨分子的細胞旁通路 (paracellular passage),將其穿越微血管 EC 內襯的途徑限制為窗孔 (fenestrae) 或囊泡運輸 (vesicular transport),從而使其能控制哪些分子可以通過(選擇性通透性)。PCs 可能透過穩定 EC–EC 接觸來影響屏障,但在皮膚微血管中過於稀疏,無法形成能直接阻擋巨分子通行的連續表面。微血管雖然直徑比其他微血管窄,但其累積表面積遠超過微血管系統的其他部分,因此大部分的通行都發生在 SVP 的這一部分。藉由允許液體與溶質經由細胞旁路徑穿越 EC 內襯,同時防止巨分子如此通行,SVP 的真皮微血管讓微血管環小動脈側附近的靜水壓 (hydrostatic pressure) 梯度驅動液體與溶質進入組織,然後在微血管環的小靜脈側,透過由保留在血管管腔內之蛋白質所形成的滲透壓 (oncotic) 梯度,將它們重新吸回血流中。微血管後小靜脈的 ECs 即使形成緊密連接也很少,而主要透過黏著連接 (adherens junctions) 維繫在一起,這些黏著連接主要由 VE-鈣黏蛋白 (VE-cadherin) 與相關的連環蛋白 (catenins) 所形成。(皮膚小動脈與微血管的 ECs 也形成黏著連接,但一旦緊密連接形成,這些黏著連接似乎是可有可無的。)在基礎狀態下,小靜脈對巨分子有些許滲漏性,但巨分子逸出至組織的情況受限於小靜脈 EC 內襯(相較於微血管而言)相對較小的表面積。這些結構特徵的淨結果是,真皮血管 ECs 是調節而非阻止血液與組織之間的分子交換。
灌流 (PERFUSION)
ECs 主要透過作用於環繞小動脈的 SMCs 來調節局部血流(圖 9-5)。這些 SMCs 的排列方向幾乎平行於血管的環周軸或徑向軸 (radial axis)。ECs 控制 SMC 張力 (tone),而 SMC 張力調節血管管腔的大小及血管對擴張的阻力。血管阻力連同血壓共同決定血流。PCs 也具有收縮性,主要功能是為終末小動脈、微血管與微血管後小靜脈提供結構完整性,但也可能對血管阻力有所貢獻。在正常情況下,ECs 同時合成血管擴張劑(vasodilators,例如一氧化氮與前列環素,可降低 SMC 張力)與血管收縮劑(vasoconstrictors,例如內皮素 endothelin,可增加張力)。EC 也表現血管收縮素轉化酶 (angiotensin-converting enzyme),可從循環中的惰性前驅物血管收縮素 I (angiotensin I) 產生血管收縮劑血管收縮素 II (angiotensin II)。血管擴張劑與血管收縮劑之間的平衡可能受神經或荷爾蒙訊號的調節。重要的是,抑制 ECs 中的一氧化氮合成酶-3 (nitric oxide synthase-3,又稱內皮型 NOS endothelial NOS) 會導致血管收縮與阻力增加,顯示血管擴張在正常情況下佔主導地位。近期證據顯示,對某些氧化脂質加合物 (oxidized lipid adducts) 具特異性的駐留血管周圍記憶 T 細胞,可透過釋放諸如腫瘤壞死因子 (tumor necrosis factor, TNF)-α 與介白素 (interleukin, IL)-17A 等細胞激素來誘導血管收縮,不過尚不清楚此現象是否發生於皮膚中。⁶
體溫調節 (THERMOREGULATION)
皮膚是熱資訊的來源,並且是熱恆定 (thermal homeostasis) 的執行器官。溫度透過初級體感覺傳入神經 (primary somatosensory afferent nerves) 來感知;皮膚血流則透過交感神經性血管擴張或血管收縮加以調節。為了保存熱量,皮膚血管收縮;為了將熱量從核心對流傳遞至周邊,皮膚血管擴張,血流增加。皮膚血流正常為 250 至 300 mL/min,但可從寒冷時的接近於零,變化至高達 6 至 8 L/min,此時約涵蓋心輸出量的 60%。⁷ 非無毛皮膚 (Nonglabrous skin) 受交感神經系統的反射性體溫調節所支配。腎上腺素性血管收縮系統 (noradrenergic vasoconstrictor system) 持續性地活躍,並由寒冷暴露所活化。對身體加熱的初始血管擴張是透過反射性地移除主動血管收縮張力來達成。隨著進一步加熱,副交感膽鹼性神經 (parasympathetic cholinergic nerves) 主要釋放乙醯膽鹼 (acetylcholine),引起主動血管擴張。同時,自律神經與感覺神經纖維釋放神經傳遞物質,例如 P 物質 (substance P)、降鈣素基因相關胜肽 (calcitonin gene-related peptide)、血管活性腸胜肽 (vasoactive intestinal peptide) 與腦下垂體腺苷酸環化酶活化多胜肽 (pituitary adenylate cyclase activating polypeptide),這些物質直接或間接地誘導血管擴張。局部因子,例如靜脈充血 (venous congestion) 與跨壁壓 (transmural pressure) 增加,也會調節皮膚血流。無毛皮膚 (Glabrous skin) 透過從動脈直接進入靜脈床、從而繞過微血管環的分流 (shunts) 來促進體溫調節,手部的熱敏感性顯著高於足底(表 9-2)。皮膚體溫調節的失調可見於與雌激素 (estrogen) 濃度改變相關的停經後熱潮紅 (postmenopausal hot flushes)。雌激素具有血管擴張功能,如在雌激素受體基因剔除 (estrogen receptor knockout) 小鼠中所證明。雌激素補充能緩解這些個體的熱潮紅,並降低其靜息體溫。第 1 型糖尿病 (Type 1 diabetes) 患者相對於健康對照組具有較高的皮膚血流,這可能與高胰島素血症 (hyperinsulinemia) 所誘導的中度血管擴張,以及皮膚血管受損的體溫調節反應有關。與受損熱反應相關的潛在機轉大多仍未獲解決。β 細胞所產生之 C-胜肽 (C-peptide) 的缺乏,或因 NOS 活性降低而導致一氧化氮生物可用性 (bioavailability) 的減少,可能扮演某種角色。一種病理性的體溫調節反射可見於雷諾氏病 (Raynaud disease);患者對寒冷或情緒有過度的血管痙攣反應 (vasospastic response),導致手指缺血 (ischemia)。雷諾氏病的血管痙攣發作侷限於供應具有豐富動靜脈吻合 (arteriovenous anastomosis) 密度之皮膚部位的皮膚動脈,即無毛部位的情況。這些部位正常情況下會在寒冷反應中受到交感腎上腺素性血管收縮活動的標靶作用。雷諾氏病患者具有增加的交感血管收縮活動,這導致通過動靜脈吻合的血流增加。值得注意的是,拮抗內皮素 A 與內皮素 B 受體 (endothelin A and endothelin B receptors) 的波生坦 (bosentan)——在硬皮症 (scleroderma) 中具有可喜的臨床效果——並不影響雷諾氏病的血管痙攣。紅斑性肢痛症 (erythromelalgia) 患者有四肢的灼痛 (burning pain)、血管擴張與紅斑。它是由影響編碼 SCN9A 基因的突變所引起,導致通道病變 (channelopathies)。雖然此異常血管反應的主要病因不明,但大多數患者的定量泌汗運動軸突反射試驗 (quantitative sudomotor axon reflex tests)(測量靜息皮膚溫度、靜息出汗量與受刺激出汗量)結果異常,有些患者有異常的腎上腺素性功能,其他患者則有異常的心臟迷走神經功能 (cardiovagal function)。紅斑性肢痛症最嚴重侵犯手掌與足底。因此,一般認為小動脈–小靜脈分流 (arteriole–venule shunts) 受到異常的神經支配,或表現出異常的反應。如先前所指出,這些小動脈–小靜脈分流主要見於無毛皮膚。
免疫監視 (IMMUNE SURVEILLANCE)
除了作為物理屏障之外,皮膚還具有免疫屏障的特徵。駐留於皮膚中的細胞之一,包括半專業抗原呈現細胞 (semiprofessional antigen-presenting cells),例如角質細胞、ECs 與巨噬細胞,以及專業抗原呈現細胞 (professional antigen-presenting cells),例如蘭格罕細胞 (Langerhans cells) 與真皮樹突狀細胞 (dermal dendritic cells)。此外,皮膚中也分佈著 T 細胞。這些皮膚 T 細胞大多為駐留性,主要位於表皮內,但如前所述,在小靜脈的血管周圍腔室中似乎也有駐留的 T 細胞。除了駐留細胞群之外,還有對歸巢至皮膚具有偏好性的循環 T 細胞。三十多年前,「皮膚相關淋巴組織 (skin-associated lymphoid tissues)」一詞被創造出來(隨後更名為皮膚免疫系統 skin immune system),它假設存在著對皮膚展現特殊親和力的 T 淋巴細胞亞群,且 T 細胞獲得此種親和力是由來自駐留皮膚細胞所接收的訊號所決定。在皮膚內的免疫反應之後,會發展出一個記憶 T 細胞亞群,它們可經由淋巴管 (lymphatics) 離開皮膚進入引流淋巴結 (draining lymph nodes)。當受到相同抗原的再次激發時,這些 T 細胞隨後可能重新進入皮膚⁸,並可能保留對皮膚部位的偏好。第一個被描述的、表現於某個記憶 T 淋巴細胞群上的皮膚相關蛋白被命名為皮膚淋巴細胞抗原 (cutaneous lymphocyte antigen, CLA)。⁹ CLA 是 T 細胞表面的一種碳水化合物部分 (carbohydrate moiety),與唾液酸化的 Lewis X (sialylated Lewis X) 密切相關,後者與 P-選擇素醣蛋白-1 (P-selectin glycoprotein-1) 的核心結構具有同源性。CLA 已被證明可結合表現於活化內皮上的 E-選擇素 (E-selectin),但後者普遍表現於活化內皮上(不過或許比在皮膚中更為短暫),顯示這並非調節皮膚特異性白血球歸巢的唯一機轉。一個提供額外特異性的潛在候選者是趨化激素 TARC(又稱為皮膚 T 細胞吸引趨化激素 cutaneous T cell-attracting chemokine 或 CCL17)(圖 9-6),它由皮膚小靜脈所產生,在發炎皮膚中被誘導,並在腸道血管上缺如。¹⁰ 其受體 CCR4 表現於 CLA 陽性的記憶 T 細胞上。TARC 與 CCR4 的交互作用觸發 CLA 陽性 T 細胞的 E-選擇素介導性黏附。然而,CCR4-/- 小鼠的皮膚浸潤並未受損,顯示存在進一步的調節途徑。另一個皮膚特異性招募 CLAhi+ T 細胞的候選者是趨化激素 CTACK(又稱為胸腺及活化調節趨化激素 thymus and activation-regulated chemokine 或 CCL27)。CTACK 由基底層角質細胞 (basal keratinocytes) 固有性表現,並可在乳頭層真皮的 ECs 與纖維母細胞 (fibroblasts) 上被偵測到。¹¹ 一般認為 CTACK 主要由表皮中的角質細胞表現與分泌,並透過醣胺聚糖 (glycosaminoglycans) 或透過其受體 CCR10,被真皮細胞捕捉與呈現(見圖 9-6)。CCR10 表現於 T 細胞上,對於它們遷移進入未發炎的皮膚至關重要。CCR10 的剔除會改變皮膚中駐留調節性 T 細胞 (regulatory T cells, Tregs) 與 CD4+ 效應 T 細胞 (effector T cells) 之間的平衡,並在致敏小鼠局部塗抹卵白蛋白 (ovalbumin) 後引起過度反應的發炎。使用中和性抗 CTACK 抗體 (neutralizing anti-CTACK antibodies),CD4/CLA/CCR10 陽性 T 細胞的歸巢會被阻斷。¹¹
循環樹突狀細胞也可能利用 CLA–E-選擇素交互作用來外滲進入皮膚。一個 B 細胞亞群也可能表現 CLA 並展現對 E-選擇素增強的結合,這可能使其能進行皮膚遷移。事實上,雖然非常罕見,B 細胞確實存在於正常人類皮膚中。¹² 這些細胞展現出一種克隆受限模式 (clonally restricted pattern),指出其辨識一種受限的抗原譜系——最可能是針對皮膚相關抗原——暗示著皮膚駐留記憶 B 細胞群存在的可能性。它們已被證明可利用 CCR6–CCL20 交互作用,從皮膚再循環至局部淋巴結。血液中也含有一個循環 T 細胞群,其特徵為效應記憶 T 細胞 (effector memory T cells),功能為尋找先前曾對其產生免疫反應的微生物抗原的存在。這些細胞與其他 T 細胞一樣,利用一套克隆分佈的受體譜系來偵測抗原,這些受體被選擇用以偵測結合於第一類主要組織相容性複合體 (class I major histocompatibility complex, MHC) 分子(CD8+ T 細胞的人類白血球抗原 human leukocyte antigen [HLA]-A、-B 或 -C)或結合於第二類 MHC 分子(HLA-DR、-DP 或 -DQ)上的微生物胜肽,但不偵測源自自體蛋白 (self-proteins) 的胜肽。真皮微血管 ECs 在基礎(未發炎)條件下即高度表現兩類 MHC 分子,並能有效地形成胜肽–MHC 分子複合體。這些細胞也表現表面蛋白,例如 LFA-3、4-1BB 配體、Ox40 配體與 ICOS 配體,這些蛋白可提供循環記憶 T 細胞活化所需、與抗原無關的增強訊號(「共刺激 costimulation」)。
ECs 在受感染組織管腔表面對抗原的辨識,不僅活化這些細胞,也觸發它們穿越 EC 內襯、滲出 (diapedesis) 進入皮膚。此過程的細胞生物學是獨特的:T 細胞對 EC 表面抗原的辨識,會使細胞變圓而非鋪展,並透過伸出一個含有微管組織中心 (microtubule organizing center) 與細胞質顆粒的細胞質突起來穿越 ECs 之間,而非讓這些結構跟隨細胞體與細胞核在後拖的尾足 (uropod) 中。ECs 的此項抗原呈現功能可能與血管周圍樹突狀細胞或巨噬細胞的抗原呈現相重複,後兩者也在突入血管管腔的延伸結構上展示胜肽–MHC 分子複合體。這兩種類型的訊號都可能補充由位於血管周圍單位內的駐留或再循環 T 細胞所啟動的免疫反應。
急性發炎的真皮血管改變 (DERMAL VASCULAR ALTERATIONS OF ACUTE INFLAMMATION)¹³
微血管系統的不同節段以不同方式促成急性發炎。歷史上,許多焦點集中在 EC 內襯所發生的變化上。局部發炎部位受到較高程度的灌流,主要是因為小動脈 ECs(以及程度較輕的微血管 ECs)所釋放之血管擴張分子的平衡發生了轉變。此變化的主要介質是前列環素合成的增加,這很可能源自細胞激素誘導的環氧合酶 (cyclooxygenase, COX)-2(又稱前列腺素 H 合成酶 2 prostaglandin H synthase 2)表現增加,COX-2 比固有性表現的 COX-1 更具活性。所造成的血流增加導致發炎部位局部溫暖(「熱 calor」)與發紅增加(「紅 rubor」)。小靜脈 EC 破壞其細胞內黏著連接(圖 9-7),允許諸如纖維連接蛋白 (fibronectin) 與纖維蛋白原等大型血漿蛋白的外滲,後者在血管外空間轉化為纖維蛋白,產生腫脹(「腫 tumor」)。這些蛋白質在組織中形成一個暫時性基質 (provisional matrix),將允許外滲的白血球附著與遷移,並將最終用於組織修復。其他變化在於小靜脈 ECs 的表面分子展示,這些分子可作用於捕捉循環白血球。EC 所表現的選擇素 (selectins) 引起白血球的繫留 (tethering) 與滾動 (rolling)。趨化激素,尤其是針對嗜中性球的 IL-8 (CXCL8) 與針對單核球的單核球趨化蛋白 1 (monocyte chemoattractant protein 1, MCP-1,又稱 CCL2),引起白血球的鋪展與牢固附著。白血球在 EC 表面朝細胞外連接處的爬行 (crawling) 主要由 ICAM (細胞間黏附分子 intercellular adhesion molecule)-1 所介導。對於單核白血球而言,血管細胞黏附分子 (vascular cellular adhesion molecule, VCAM)-1 可能有助於捕捉、滾動與爬行。趨化激素可能由 ECs 製造,或可能由血管周圍單位的哨兵細胞所產生,並透過非共價地附著於 EC 管腔表面的肝素硫酸鹽 (heparin sulfates) 而被展示出來。ECs 的這些變化被稱為內皮活化 (endothelial activation)。某些自泌素 (autacoids) 或酵素(例如凝血酶)可誘導短暫的、與蛋白質合成無關的變化,例如 P-選擇素從細胞內儲存顆粒(稱為 Weibel-Palade 小體 Weibel-Palade bodies)轉位至細胞表面,或合成趨化性脂質,例如血小板活化因子 (platelet activating factor)。此類變化有時被稱為第 I 型活化 (Type I activation),但此類變化的短暫性質通常導致短期效應,例如風疹塊與潮紅反應 (wheal and flare reaction),而非白血球招募。後者更依賴於由發炎細胞激素(例如 TNF-α、IL-1α 或 IL-1β)所誘導的新基因轉錄與蛋白質合成。EC 的細胞激素活化也可透過下調血栓調節蛋白合成、同時誘導組織因子的表現來促進凝血。凝血可能透過 EC 釋放含有組織因子之質膜衍生微粒 (plasma membrane-derived microparticles) 而傳播。當白血球到達微血管後小靜脈 ECs 的細胞內連接處時,它們會接合促成外滲的額外蛋白質。其中數種,例如血小板內皮細胞黏附分子 (platelet endothelial cell adhesion molecule, PECAM)-1、小兒麻痺病毒受體 (poliovirus receptor) 與 CD99,儲存於側邊緣質膜的內陷處(稱為側邊緣再循環腔室 lateral border recycling compartment),它們被帶到細胞表面以接合白血球上所表現的受體。(有時這些分子可排列於位於實際細胞間連接附近、但不在其上的短暫跨細胞通道 transcellular channels,允許白血球穿過 ECs 而非從 ECs 之間通過。尚不清楚此種區別是否具有任何生理意義。)在白血球穿過 EC 層之後,它接合基底膜蛋白與 PCs,後者因應與活化 ECs 相同的細胞激素而表現升高濃度的 ICAM-1 與趨化激素。隨著 PCs 收縮以打開基底膜中的縫隙並引導外滲白血球至這些區域,與 PCs 的交互作用可能延續 30 分鐘或更久。當位於基底膜之外時,外滲的白血球繼續與 PCs、或許還有血管周圍單位的其他細胞交互作用,沿著血管的外膜表面遷移。TNF-α 與 IL-1 在 ECs 中誘導促發炎變化,這些變化通常不區分不同的白血球類型。然而,不同黏附分子的誘導動力學各不相同。E-選擇素在最初 2 小時內製造,並與嗜中性球招募的開始相關,而 VCAM-1 較晚合成(即約 6 至 12 小時),並與單核白血球招募的開始相關(圖 9-8)。極化細胞激素 (Polarizing cytokines),例如干擾素 (interferon, IFN)-γ 或 IL-4 與 IL-13,分別有利於產生優先活化 T 細胞與效應細胞的趨化激素,這些細胞大多與 TH1 或 TH2 型後天免疫相關。IL-17 對 ECs 影響甚微,反而作用於 PCs,誘導延長嗜中性球存活的因子(顆粒球-巨噬細胞群落刺激因子 granulocyte-macrophage colony-stimulating factor 與顆粒球群落刺激因子 granulocyte-colony stimulating factor),有利於 TH17 型發炎所特有之嗜中性球發炎的延長。¹⁴
慢性發炎的真皮血管改變 (DERMAL VASCULAR ALTERATIONS OF CHRONIC INFLAMMATION)
ECs 的持續活化是可能的,並導致與急性活化相比截然不同的變化。以內皮特異性方式過度表現 TNF 的基因轉殖小鼠展現出一種發炎性皮膚表型,伴隨表皮過度增生 (epidermal hyperproliferation) 與纖維化 (fibrosis)。在培養中,慢性 TNF 刺激的 EC 雖然展現持續且高量的 ICAM-1 與 VCAM-1 表現,但 E-選擇素缺如,且無法藉由額外的 TNF 刺激重新誘導(即 ECs 對 E-選擇素表現產生減敏化 desensitized)。有趣的是,這些慢性 TNF 刺激的細胞持續過度表現 CXCL5 與 MCP-3。¹⁵ 與臍靜脈內皮 (umbilical vein endothelium) 相比,培養的皮膚 ECs 顯示出持續且較高的 E-選擇素表現,以及較慢的 E-選擇素蛋白內化 (internalization) 與降解,但這可能是微血管 ECs 的一般特性。¹⁶ 儘管如此,E-選擇素的持續表現雖然並非皮膚所獨有,但一般被描述為慢性發炎的皮膚特異性特徵(見圖 9-8)。
特定疾病中的真皮血管變化 (DERMAL VASCULAR CHANGES IN SPECIFIC DISEASES)
急性與慢性皮膚發炎:一般考量 (ACUTE VERSUS CHRONIC SKIN INFLAMMATION: GENERAL CONSIDERATIONS)
在急性皮膚炎中,ECs 表現高量的 E-選擇素與 P-選擇素,以及中等量的 VCAM-1 與 ICAM-1。發炎浸潤主要由 CLA 與 CD45RO 陽性的記憶 T 細胞所組成。在慢性皮膚病灶中,例如乾癬 (psoriasis)、慢性紅斑性狼瘡 (chronic lupus erythematosus) 與蕈狀肉芽腫 (mycosis fungoides),ECs 表現中等量的 E-選擇素與高量的 VCAM-1。它們也可能經歷型態變化,並獲得一種令人聯想到高內皮小靜脈 (high endothelial venules) 的表型。此類 EC 具有立方狀 (cuboidal) 外觀,不同於內襯其他血管之 ECs 的扁平型態。高內皮小靜脈表現唾液酸化、岩藻糖化與硫酸化的碳水化合物部分,對應於周邊淋巴結地址素 (peripheral node addressins),這些可被抗體 MECA-79 所辨識。這些地址素 (addressins) 固有性地表現於淋巴結的高內皮小靜脈上;它們結合於白血球所表現的 L-選擇素 (L-selectin),並控制 T 細胞通過淋巴結的再循環。這些唾液酸化、岩藻糖化與硫酸化的碳水化合物部分也可能表現於慢性皮膚病灶中,例如皮膚 T 細胞淋巴瘤 (cutaneous T-cell lymphoma)。¹⁷ 在皮膚黑色素瘤 (cutaneous melanoma) 中,血管也表現 MECA-79 部分,且 MECA-79 反應性血管的數量與腫瘤浸潤淋巴細胞 (tumor infiltrating lymphocytes) 的數量及腫瘤消退 (tumor regression) 的組織學徵象相關。¹⁸ 在鱗狀細胞癌 (squamous cell carcinoma) 中,MECA-79 陽性血管的數量被發現與存活率相關。
乾癬 (PSORIASIS)
如前所述,白血球主要透過微血管後小靜脈離開血流。真皮乳頭尖端內的微血管環具有小動脈型表型 (arteriolar phenotype);因此,白血球可透過下方的 SVP 離開血流,但無法在乳頭尖端內離開。在乾癬中,情況有所不同。在此疾病中,微血管環延長並改變型態。正常微血管在迴轉點 (turnaround point) 坐落於小動脈型基底膜上,而乾癬性微血管環在迴轉點則接觸表皮,此現象稱為「親吻 (kissing)」,並重塑其基底膜使其類似於微血管後小靜脈的基底膜。環內的 ECs 增生,同時產生延長與直徑的增加。(ECs 的增生有時被誤解為血管新生,但實際上並未形成新血管。)此外,重塑後的乾癬性環的 ECs 表現通常與細胞激素活化之小靜脈 ECs 相關的黏附分子,支持白血球的繫留、滾動與外滲。¹⁹ 淋巴細胞與嗜中性球現在可以在乳頭尖端內離開血管管腔並遷移進入表皮。這種淋巴細胞與嗜中性球在乳頭尖端內外滲的組織學圖像被稱為「噴射狀乳頭 (squirting papillae)」,是乾癬的典型特徵,並與慢性濕疹 (chronic eczema) 的發現形成對比——在慢性濕疹中,白血球穿越 SVP 的微血管後小靜脈外滲(圖 9-9)。
紅色扁平苔癬 (LICHEN RUBER PLANUS)
另一種慢性皮膚疾病——紅色扁平苔癬 (lichen ruber planus)——與健康皮膚相比,在乳頭層真皮內具有擴大的血液微循環床。具體而言,口腔扁平苔癬 (oral lichen planus) 具有彎曲的微血管,並伴隨微血管環直徑的擴大。在一小系列患者中,病灶內注射貝伐單抗 (bevacizumab,抗 VEGF 抗體) 誘導病灶消退,且在 3 個月的追蹤期間無復發。在此情境下,血管內皮生長因子 (vascular endothelial growth factor, VEGF) 不僅誘導血管新生,也透過誘導內皮黏附分子表現而具有促發炎性,這可能解釋抗 VEGF 抗體的效果。
皮膚血管系統在傷口癒合與癌症中的角色 (SKIN VASCULATURE IN WOUND HEALING AND CANCER)
血管新生與血管生成 (ANGIOGENESIS AND VASCULOGENESIS)
在健康成人中,血管是穩定的結構,ECs 的更替 (turnover) 非常緩慢。在慢性發炎、組織損傷或腫瘤生長的情境中,可能形成新血管,且既有血管可能經歷重塑。血管形成過程的大部分知識,是透過研究胚胎發育 (embryogenesis) 期間血管系統的形成而獲得的。在胚胎中,ECs 起源於血管母細胞 (angioblasts),後者遷移至周邊組織並形成原始血管(血管生成 vasculogenesis)。隨後,新血管從既有血管中產生(血管新生 angiogenesis)(圖 9-10)。間葉細胞 (Mesenchymal cells) 接著被招募進入血管壁,並在其中後續分化為 SMCs 與 PCs,此過程稱為血管重塑 (vascular remodeling)。在成人中,新血管透過血管新生(從既有血管萌芽 sprouting)而發展;有人提出循環內皮前驅細胞 (circulating endothelial progenitor cells)、中胚層血管母細胞 (mesoangioblasts) 與多能性成人前驅細胞 (multipotent adult progenitor cells) 促成新血管形成(成人血管生成 adult vasculogenesis)。數位研究者提出這些前驅細胞起源於骨髓,正如血管母細胞起源於胚胎的血島 (blood island)。然而,骨髓衍生細胞是否能產生穩定的 ECs 仍不清楚,而許多此類觀察現今已被重新詮釋為暗示著表現 EC 標記、促進血管新生、然後在新血管變得穩定時消失的單核球所扮演的角色。幹細胞 (Stem cells) 與內皮前驅細胞已在血管壁中被辨識為駐留細胞。此類細胞被認為駐留於一個微環境 (niche) 中,等待適當的刺激(例如血管損傷)以參與受損血管的修復與新血管的形成。在此類血管壁前驅細胞中,有分化為 ECs 的 CD34+/CD31- 細胞。其他駐留血管壁前驅細胞為 SMC 前驅細胞與可能源自 PCs 的間葉基質細胞 (mesenchymal stromal cells)。
皮膚中誘導新血管形成的訊號遵循所有器官中所給定的主要機轉。VEGF 家族的分子——VEGF A 至 E——最顯著地作用於 ECs。纖維母細胞生長因子 (Fibroblast growth factor, FGF) 1 與 2 也是強效的 EC 促有絲分裂劑 (mitogens),但也作用於 SMCs、PCs 與纖維母細胞。在成人中,FGF-1 與 FGF-2 儲存於各種細胞的細胞質中,並在細胞受損時釋放,作為「傷口荷爾蒙 (wound hormones)」以刺激局部血管新生與結締組織生長。其他多胜肽因子也已被發現在體內與體外作用於 ECs,包括表皮生長因子 (epidermal growth factor, EGF)、肝素結合 EGF 樣生長因子 (heparin-binding EGF–like growth factor),以及肝細胞生長因子 (hepatocyte growth factor, HGF) 或散佈因子 (scatter factor, SF)。對於血管重塑與穩定化,需要酪胺酸激酶受體 1 (tyrosine kinase receptor 1, Tie) 與 2。Tie-2 透過結合血管生成素 (angiopoietins),介導間葉細胞與未成熟血管內皮之間的對話(圖 9-11)。eph 家族的受體酪胺酸激酶 (receptor tyrosine kinases) 與其對應的 ephrin 配體是膜結合性的,且似乎介導雙向的細胞間訊號傳遞。在血管發育期間,ephrin-B2 標記早期動脈血管的 ECs,而 ephrin-B4 標記早期靜脈的 ECs。血小板衍生生長因子 (Platelet-derived growth factor, PDGF) 與轉化生長因子 (transforming growth factor, TGF)-β 由 ECs 分泌,並促進間葉細胞的遷移、基質生物合成與分化。基質醣蛋白 (Matrix glycoproteins) 例如纖維連接蛋白與層黏連蛋白 (laminin),以及基質醣蛋白的受體例如 β1 與 β3 整合素 (integrins),也被認為在血管生成與血管新生中扮演角色。這似乎符合直覺,因為除非 ECs 與間葉細胞透過整合素受體與基質蛋白交互作用,否則它們都無法存活。
傷口癒合 (WOUND HEALING)
全層傷口 (Full-thickness wounds) 透過相互連接且重疊的階段癒合,涉及凝血、發炎、增生與重塑。受傷後立即,被破壞的血管收縮以止血。凝血級聯 (coagulation cascade) 的活化導致血小板血塊,並隨後形成纖維蛋白血塊。在此階段,受傷的血管作為循環發炎細胞的引導結構。補體活化 (Complement activation) 以及血小板與角質細胞釋放 CXC 趨化激素,導致循環嗜中性球的招募。周圍微血管變得有滲漏性,導致更多血漿與細胞在傷口內積聚。血管滲漏性逐漸減少,但會持續接下來的 3 至 7 天。微血管網絡的形成透過以下方式發生:從母血管 (parent vessel) 的微血管尖端延伸、微血管尖端成熟為微血管萌芽 (capillary sprouts)、吻合,以及進一步的分支。在癒合的發炎期與增生期這種血管的快速形成之後,血管系統會被重塑、退化,或兩者皆有。血管新生反應主要由 VEGF、FGF-2、PDGF 與 TGF-β 家族成員所驅動(圖 9-12)。巨噬細胞是癒合早期階段 VEGF 的主要來源;在較晚期階段,大多數表現 VEGF 的細胞位於新生上皮 (neoepithelium) 內。²⁰ 在損傷、缺氧 (hypoxia) 或發炎的條件下,角質細胞產生並釋放廣泛的血管新生因子,包括 FGF 或 TGF 蛋白家族成員、PDGF 或 VEGF。同時,VEGFR-2 在 ECs 上被上調,這增強了內皮對 VEGF 的反應。FGF-2 來自浸潤的巨噬細胞,但也來自 ECs 本身。抗 FGF-2 抗體幾乎完全抑制傷口的血管新生。來自皮膚幹細胞微環境 (skin stem cell niches) 的幹細胞與微環境之外的內源性幹細胞支持傷口癒合。這些包括毛囊隆突 (hair follicle bulge) 與環繞毛囊的真皮鞘 (dermal sheath) 幹細胞、表皮幹細胞,以及骨髓衍生的間葉幹細胞。儘管有所有的臨床前傷口癒合研究,基於幹細胞的策略在臨床情境中的轉譯僅獲得有限的成功。²¹ 與透過局部萌芽之傳統血管新生相比,幹細胞衍生之血管新生的定量貢獻仍不清楚。
腫瘤血管新生 (TUMOR ANGIOGENESIS)
皮膚腫瘤與任何其他實體腫瘤一樣,從鄰近的血液微血管獲取其血液供應。沒有足夠血液供應的快速腫瘤生長會誘導缺氧。缺氧因腫瘤血管的滲漏性與紊亂性而加劇,導致氧張力 (oxygen tension) 的急性波動以及腫瘤內擴散受限的區域 (diffusion-limited regions)。腫瘤與基質 (stroma) 兩個腔室都透過表現缺氧誘導因子 (hypoxia-inducible factor, HIF) 家族的轉錄因子來對缺氧做出反應。²² 缺氧刺激腫瘤細胞與巨噬細胞分泌 TGF-β、PDGF、CXCL2 與內皮素,這些活化纖維母細胞。在纖維母細胞中,缺氧刺激細胞外基質重塑與血管新生因子的釋放。缺氧透過降低 PCs 與 ECs 之間的關聯,直接影響血管屏障功能。腫瘤本身可能產生 VEGF、FGF-2、EGF 與 HGF 或 SF(見圖 9-12)。此外,腫瘤可能透過套疊式血管新生(intussusceptive angiogenesis,將既有血管一分為二)、血管擬態(vasculogenic mimicry,由腫瘤細胞形成導流液體的通道)、血管共選(vessel co-option,腫瘤細胞沿著既有血管遷移)與血管生成來增加血液供應。²³
血管擬態 (Vascular mimicry) 似乎在轉移性黑色素瘤 (metastatic melanoma) 中扮演特定角色(而非在原發病灶中),因為具有此類血管通道形成的患者具有較差的 5 年存活率。²⁴ 在腫瘤相關血管系統中所觀察到的另一個有趣現象是內皮-間葉轉化 (endothelial-to-mesenchymal transition),它產生癌症相關的肌纖維母細胞 (cancer-associated myofibroblasts),後者刺激發炎與纖維化、加劇血管功能障礙,並因此增加缺氧。TGF-β 是內皮-間葉轉化的驅動因子之一。²⁵
結論 (CONCLUSIONS)
皮膚血管系統在數個方面是獨特的。它被組織為功能上截然不同的血管節段:乳頭尖端內的環、SVP 與 DVP。這些節段可單獨或共同對外源性或內源性刺激產生反應,從而影響皮膚疾病的表現。由於其與表皮的鄰近性,SVP 的 ECs 可能直接對角質細胞衍生的因子(例如皮膚 T 細胞吸引趨化激素或 VEGF)做出反應,這導致血管結構、表型與功能的改變。此外,皮膚 EC 可能直接暴露於環境抗原。在此情境下,微血管後小靜脈對 HLA 第二類分子的固有表現暗示著 ECs 在抗原呈現中扮演角色,這可能增加抗原被及時辨識的可能性,從而有效率地將記憶 T 細胞招募至危險部位。
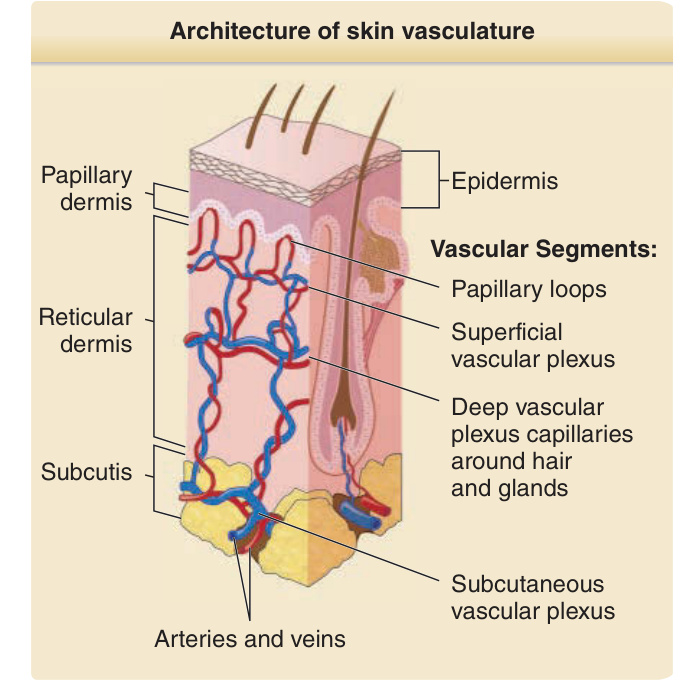
圖 9-1:皮膚血管系統結構的示意圖 (Schematic diagram of the architecture of the skin vasculature)。
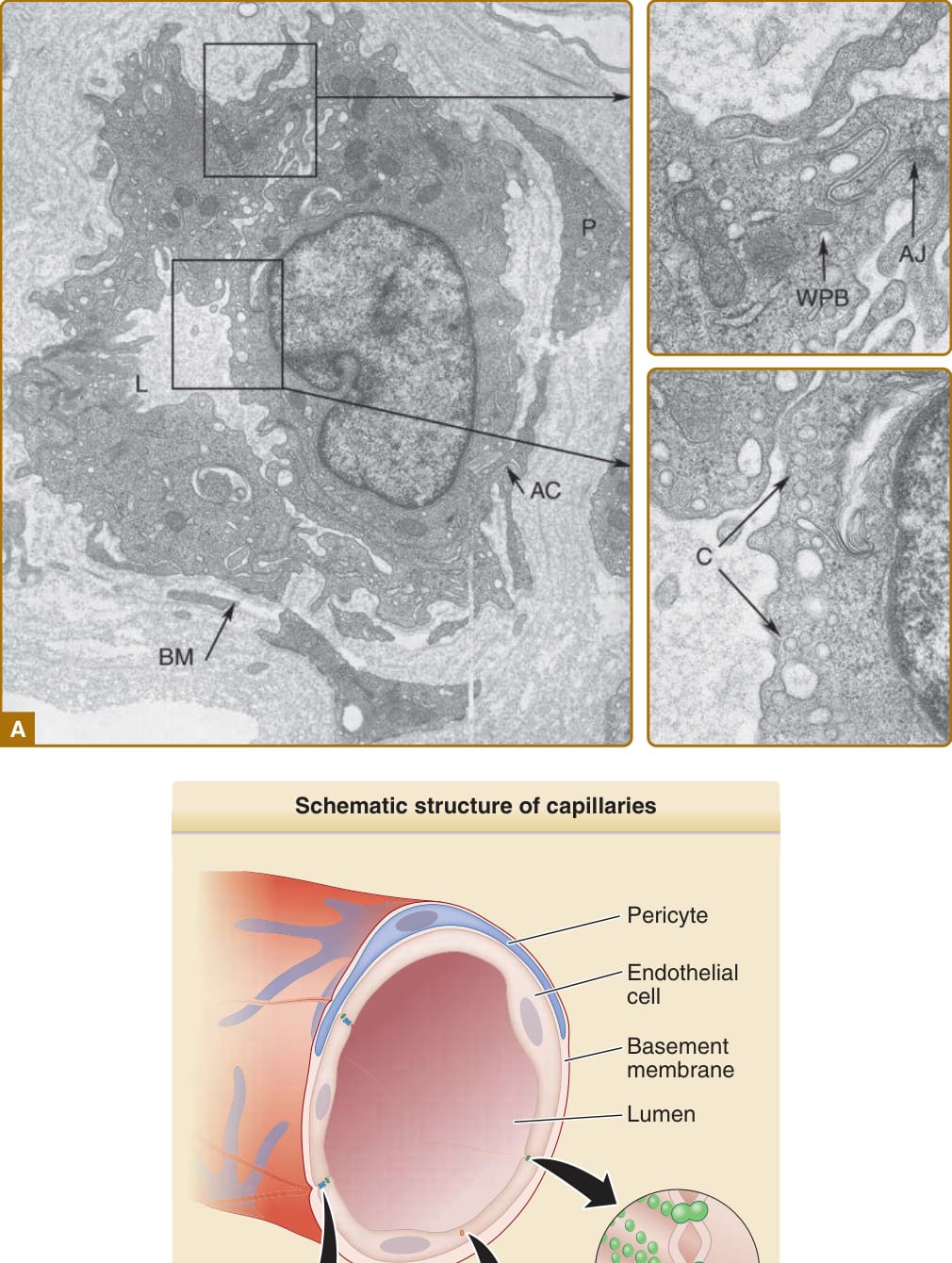
圖 9-2:A,微血管後小靜脈 (postcapillary venule) 的超微結構。AC,內皮細胞 (endothelial cell) 與周細胞 (pericyte) 之間的接觸區域;AJ,黏著連接 (adherens junction);BM,基底膜 (basement membrane);C,小窩 (caveolae);L,管腔 (lumen);P,周細胞 (pericyte);WPB,Weibel-Palade 小體 (Weibel-Palade body)。B,A 的示意圖。
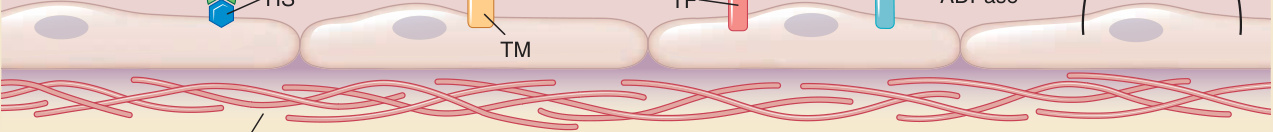
圖 9-3:內皮細胞的抗凝血與血栓功能 (Anticoagulant and thrombotic functions of endothelial cells)。為了防止凝血,內皮細胞 (1) 表現抗凝血肝素硫酸鹽 (heparan sulfates, HS),可結合並活化抗凝血酶 III (antithrombin III, ATIII),後者隨即阻斷因子 Xa(將凝血酶原 prothrombin 轉化為凝血酶)與凝血酶(將纖維蛋白原 fibrinogen 轉化為纖維蛋白 fibrin)的酵素活性;(2) 表現血栓調節蛋白 (thrombomodulin, TM),可結合凝血酶並將其從一種切割纖維蛋白原為纖維蛋白的促凝血酵素,改變為一種活化蛋白質 C (protein C) 的抗凝血酵素,蛋白質 C 與蛋白質 S (protein S) 結合後,隨即切割並去活化因子 V 與 VIII(分別作為因子 Xa 與 IXa 的輔因子);以及 (3) 表現組織因子途徑抑制因子 (tissue factor pathway inhibitor),可防止組織因子加速因子 VIIa 的催化活性,後者切割並活化因子 IX 與 X 成為因子 IXa 與 Xa。為了防止血小板活化,內皮細胞 (1) 表現外酶 (ectoenzymes),可將作為血小板活化劑的細胞外三磷酸腺苷 (adenosine triphosphate, ATP) 與二磷酸腺苷 (adenosine diphosphate, ADP) 轉化為無法活化血小板的單磷酸腺苷 (adenosine monophosphate, AMP);(2) 合成並釋放血小板活化抑制因子,例如一氧化氮 (nitric oxide, NO) 與前列環素 (prostacyclin, PGI2);以及 (3) 防止血小板與基底膜膠原蛋白(另一種血小板活性的活化劑)接觸。這些功能彼此強化,因為凝血酶是血小板的活化劑,而活化的血小板提供結合凝血因子的脂質表面以促進凝血級聯。
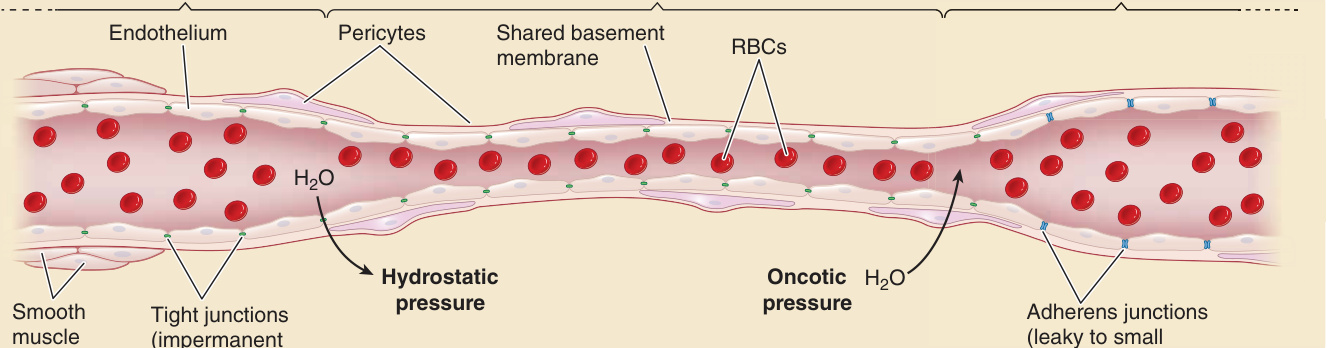
圖 9-4:內皮細胞建立選擇性通透屏障 (Endothelial cells establish permselective barriers)。選擇性通透性 (Permselectivity) 指的是微血管系統的內皮內襯允許水與溶質通過、但不允許巨分子通過的能力。表現於小動脈與微血管中的緊密連接 (tight junctions, TJs) 允許水與溶質通過,但防止巨分子與細胞通過。某些小分子(而非其他)可基於大小與電荷而通過。微血管小動脈端的靜水壓 (hydrostatic pressure) 迫使水與溶質穿越 TJs,而被保留的巨分子則在微血管小靜脈端透過滲透壓 (oncotic pressure) 將水與小分子吸回。微血管後小靜脈缺乏 TJs,其黏著連接 (adherens junctions) 受到動態調節,以允許某些巨分子在發炎情境下進入組織。RBC,紅血球 (red blood cell)。
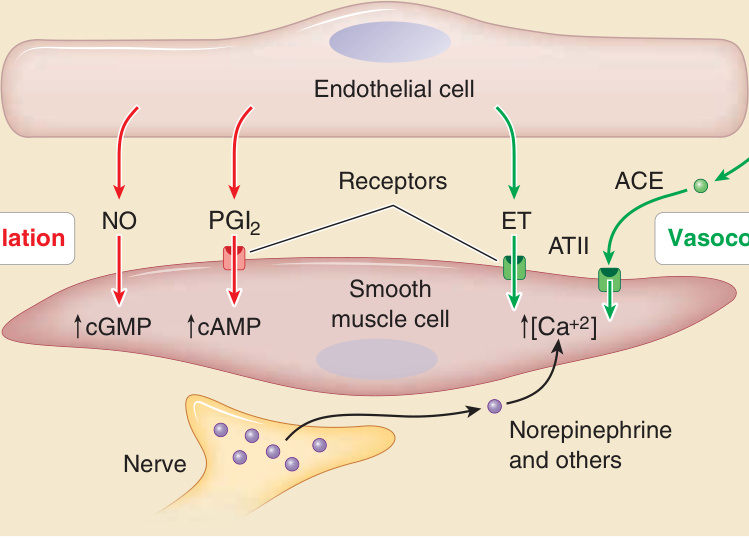
圖 9-5:透過調節平滑肌細胞張力控制灌流 (Control of perfusion by regulating smooth muscle cell tone)。血管的主動阻力可透過改變肌肉動脈與小動脈中壁性血管平滑肌細胞的收縮張力來調節。內皮細胞可產生血管擴張劑,例如一氧化氮 (nitric oxide, NO) 與前列環素 (prostacyclin, PGI2)。NO 直接活化可溶性鳥苷酸環化酶 (soluble guanylate cyclase) 以提升環狀單磷酸鳥苷 (cyclic guanosine monophosphate, cGMP) 濃度,而 PGI2 與一個膜 G 蛋白偶聯受體 (G–protein coupled receptor) 交互作用,該受體活化腺苷酸環化酶 (adenylate cyclase) 以增加環狀單磷酸腺苷 (cyclic adenosine monophosphate, cAMP)。兩者都減少肌動肌凝蛋白 (actomyosin) 收縮的程度。內皮細胞也可產生血管收縮劑,例如內皮素 (endothelin, ET),或透過外酶血管收縮素轉化酶 (angiotensin-converting enzyme, ACE),將無活性的血管收縮素 I (angiotensin I, ATI) 轉化為血管活性的血管收縮素 II (angiotensin II, ATII)。ET 與 ATII 都透過 G 蛋白偶聯受體傳訊,以增加胞質游離鈣離子的濃度,增加肌動肌凝蛋白收縮的程度。神經可能釋放神經傳遞物質,這些物質也結合於 G 蛋白偶聯受體以增加胞質游離鈣與肌動肌凝蛋白收縮。增加的肌動肌凝蛋白收縮縮短了呈放射狀排列之平滑肌細胞的長度,從而減少血管管腔直徑並增加血流阻力。
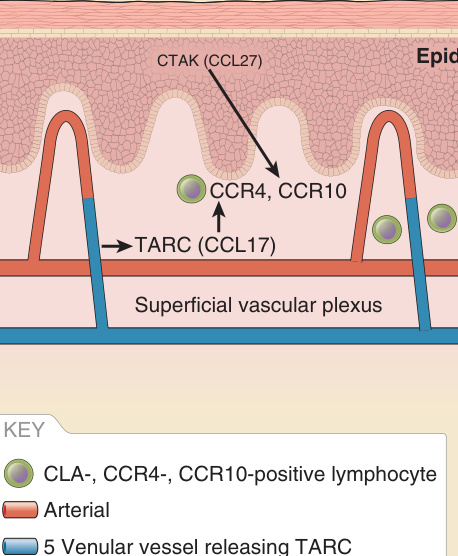
圖 9-6:免疫監視 (Immune surveillance)。
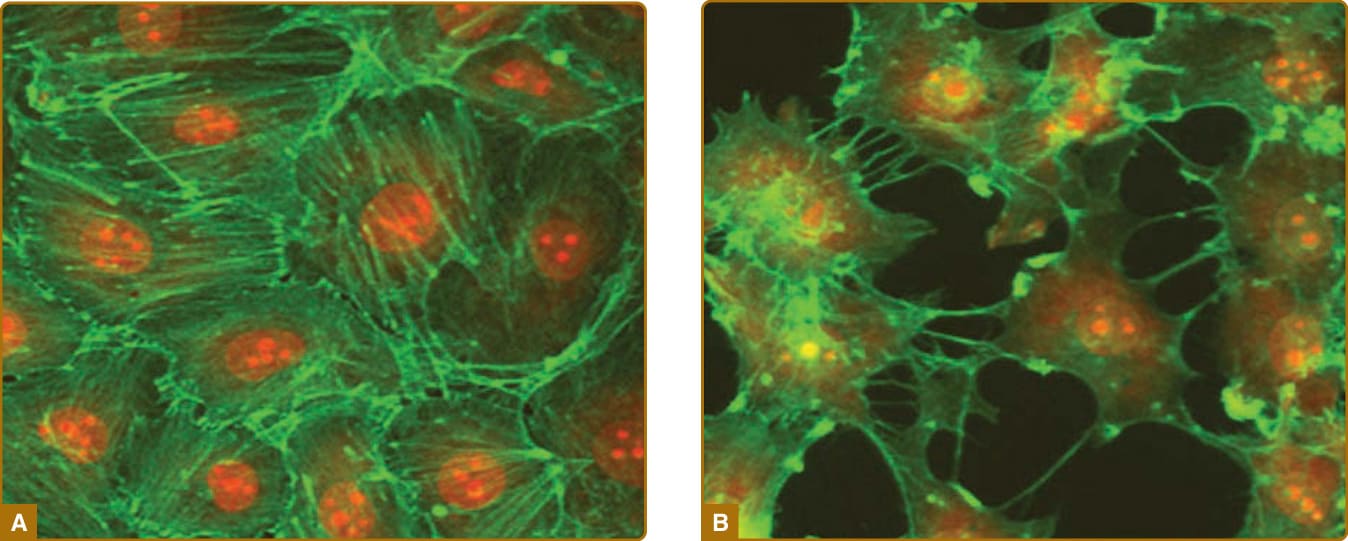
圖 9-7:培養的小靜脈內皮細胞形成連續的單層 (A);經凝血酶 (thrombin) 處理、細胞間連接被破壞而模擬滲漏血管的內皮細胞 (B)。肌動蛋白骨架 (actin skeleton) 的鬼筆環肽 (phalloidin) 染色呈綠色,細胞核呈紅色;1000× 放大倍率。
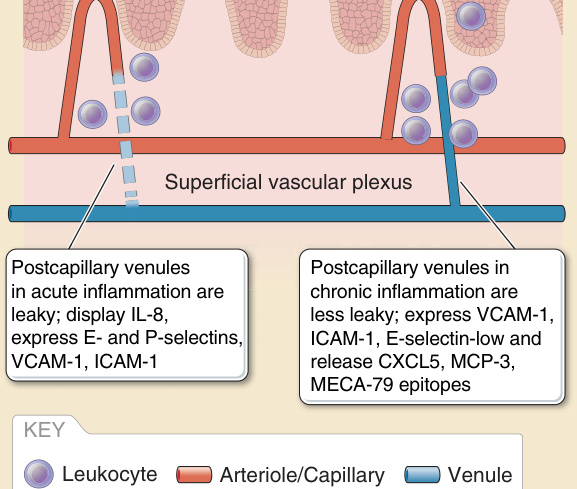
圖 9-8:急性與慢性發炎中的血管變化 (Vascular changes in acute and chronic inflammation)。ICAM,細胞間黏附分子 (intercellular adhesion molecule);IL,介白素 (interleukin);MCP-3,單核球趨化蛋白 3 (monocyte chemoattractant protein 3);VCAM,血管細胞黏附分子 (vascular cellular adhesion molecule)。
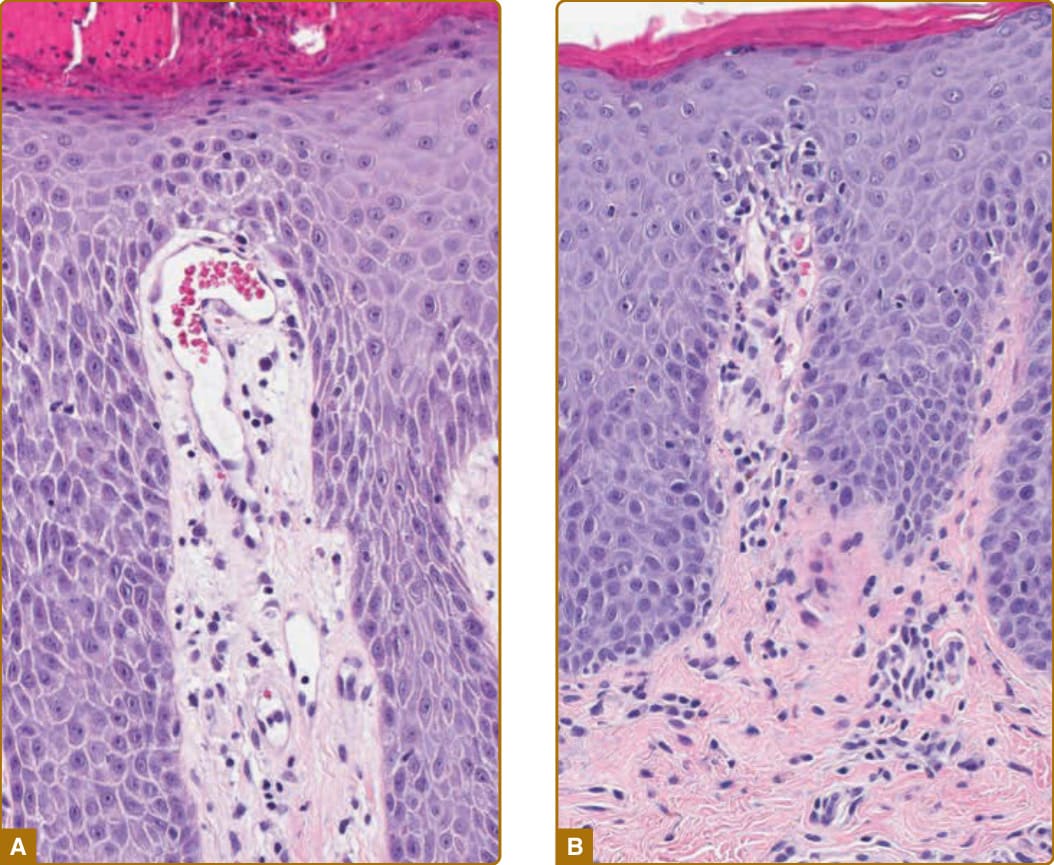
圖 9-9:A,乾癬 (psoriasis) 中的血管變化;乳頭尖端內的微血管環擴張、延長,並在迴轉點接觸表皮,此現象稱為「親吻 (kissing)」。B,白血球在乳頭尖端內離開血管管腔(「噴射狀乳頭 squirting papillae」);20× 放大倍率。
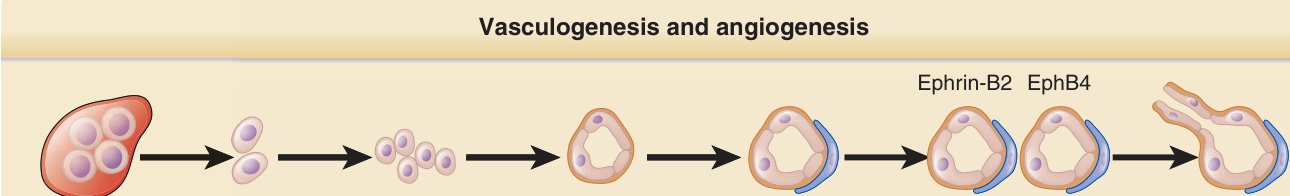
圖 9-10:血管生成與血管新生 (Vasculogenesis and angiogenesis)。ANGPT,血管生成素 (angiopoietin);FGF,纖維母細胞生長因子 (fibroblast growth factor);TGF-β,轉化生長因子-β (transforming growth factor-β);VEGF,血管內皮生長因子 (vascular endothelial growth factor)。
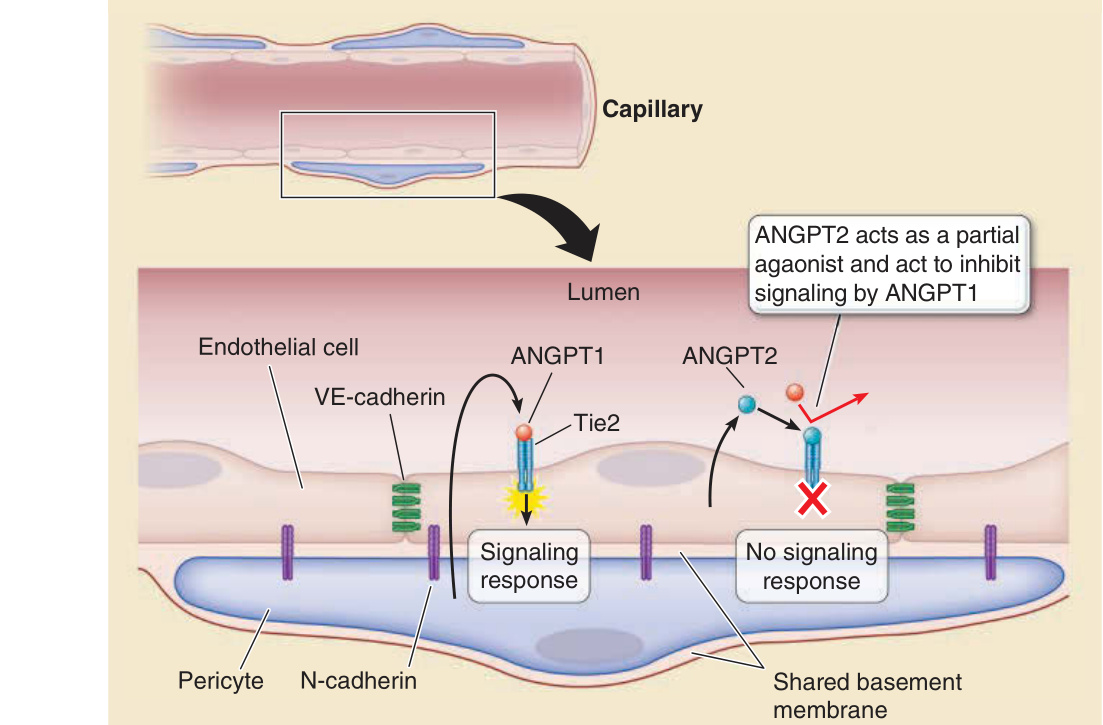
圖 9-11:內皮細胞 (endothelial cell, EC)–周細胞 (pericyte) 交互作用。周細胞定位於微血管 EC 內襯的基底膜內,並促成其合成與維持,而基底膜的組成與組織可能改變 EC 的功能。周細胞直接接觸 ECs,形成涉及 N-鈣黏蛋白 (N-cadherin) 連結的黏著連接。周細胞也透過旁分泌訊號 (paracrine signals) 穩定 ECs;其中最被了解的涉及周細胞分泌血管生成素 1 (angiopoietin 1, ANGPT1),後者結合並活化表現於內皮上的受體酪胺酸激酶 Tie2。ANGPT1 的結合防止 EC 衍生的 ANGPT2 結合到同一受體。ANGPT2 作為部分促效劑 (partial agonist),當其濃度增加時,會防止 ANGPT1 介導的訊號傳遞,導致內皮細胞連接的破壞,並與微血管滲漏、血管新生,或兩者皆有相關。
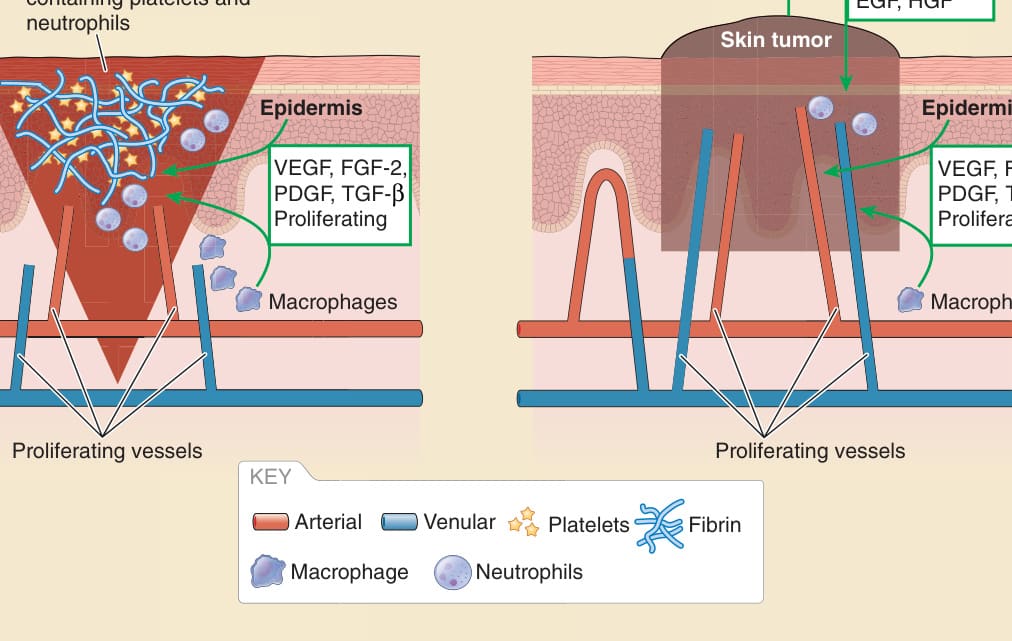
圖 9-12:傷口癒合與癌症中的血管系統 (Vasculature in wound healing and cancer)。EGF,內皮生長因子 (endothelial growth factor);FGF,纖維母細胞生長因子 (fibroblast growth factor);HGF,肝細胞生長因子 (hepatocyte growth factor);PDGF,血小板衍生生長因子 (platelet-derived growth factor);TGF-β,轉化生長因子-β (transforming growth factor-β);VEGF,血管內皮生長因子 (vascular endothelial growth factor)。

表 9-1:正常皮膚血管系統的特性與功能 (Properties and Functions of Normal Skin Vasculature)

表 9-2:非無毛皮膚與無毛皮膚的體溫調節 (Thermoregulation in Nonglabrous and Glabrous Skin)