色素沉著與黑色素細胞生物學 (Pigmentation and Melanocyte Biology)
重點一覽
- 黑色素細胞 (melanocytes) 源自神經嵴 (neural crest) 譜系;其關鍵功能是合成黑色素 (melanin) 並轉移至周圍角質細胞 (keratinocytes)。
- 可產生棕/黑色真黑色素 (eumelanin) 或橙/黃色褐黑色素 (pheomelanin);黑色素體 (melanosome) 是一種溶酶體相關胞器 (lysosome-related organelle),為黑色素合成與儲存場所。
- 對發育、遷移、存活重要的轉錄因子(MITF、SOX10、PAX3)與訊號途徑(KIT、EDNRB)在黑色素瘤 (melanoma) 中也扮演關鍵角色。
- 黑色素皮質素 1 受體 (melanocortin 1 receptor, MC1R) 常見變異造成「紅髮表型 (red hair phenotype)」——紅髮、白皙皮膚、無法曬黑、黑色素瘤風險增加;MC1R 經由 MITF 調節色素沉著與曬黑。
解剖、組織學與分布
-
黑色素細胞為樹突狀 (dendritic) 合成黑色素細胞,位於表皮基底層、毛球 (hair bulb) 與毛囊外根鞘;數目約為角質細胞的十分之一。
-
不同種族間黑色素細胞數目相似;色素差異源自黑色素的「量」與「質」,主要由角質細胞內黑色素體的數目、大小、分布與功能決定。
-
染色與標記:銀染色(Fontana-Mason、Warthin-Starry)顯示黑色素(與真黑色素性黑色素反應,褐黑色素性不反應);免疫標記有酪胺酸酶 (tyrosinase)、MITF、Melan-A/Mart1;S100 高敏感但較不特異;HMB-45(gp100/PMEL17)對黑色素細胞腫瘤提供特異性,正常成人黑色素細胞為陰性。
-
超微結構:黑色素細胞無橋粒 (desmosomes) 與張力絲;關鍵特徵為黑色素體。真黑色素體 (eumelanosomes) 依成熟度分為第 I–IV 期(I、II 期未黑色素化,III 期沿纖維沉積色素,IV 期完全黑色素化);褐黑色素體較小、各期皆呈圓形、纖維網絡較紊亂。
-
遺傳性色素疾病(與發育/生物學基因突變相關):
- 低色素:眼白化症 (OA1, GPR143)、眼皮膚白化症(OCA1 TYR、OCA2 OCA2、OCA3 TYRP1、OCA4 SLC45A2/MATP)、Waardenburg 症候群(WS1/3 PAX3、WS2 MITF/SNAI2、WS4 SOX10/EDN3/EDNRB 併赫希施普龍病)、Tietz 症候群 (MITF)、局部白斑 (piebaldism, cKIT)、Hermansky-Pudlak 症候群(黑色素體功能障礙+出血體質+肺纖維化等)、Chediak-Higashi 症候群 (LYST)、Griscelli 症候群(黑色素結塊;GS1 MYO5A、GS2 RAB27A、GS3 MLPH)。
- 高色素:家族性進行性高色素沉著 (FPH, KITLG)、家族性進行性高與低色素沉著 (FPPH, KITLG)。
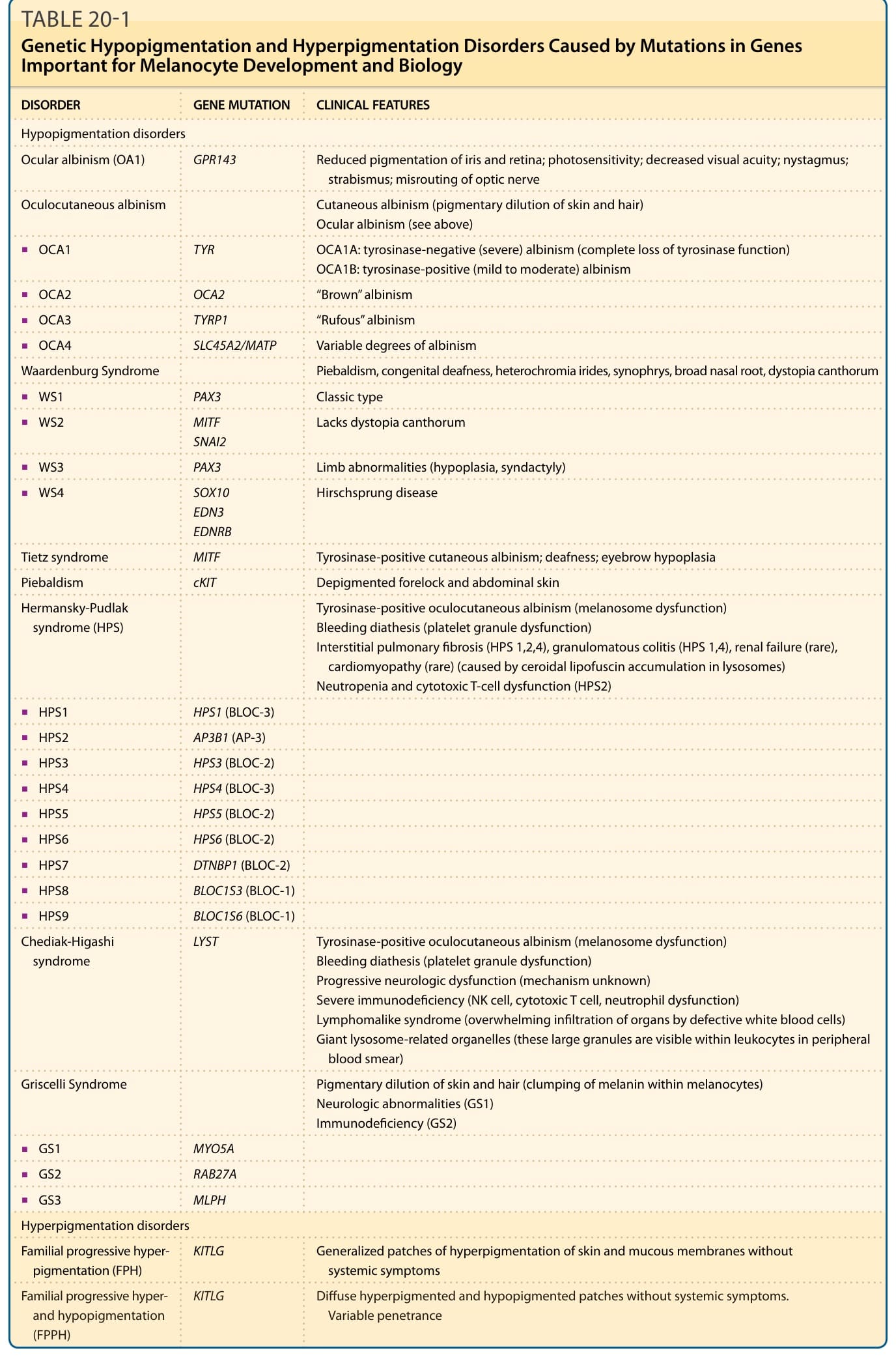
表 20-1:與黑色素細胞發育及生物學重要基因突變相關之遺傳性低色素與高色素疾病
- 分布:
- 表皮黑色素單位 (epidermal melanin unit):一個黑色素細胞透過樹突接觸並輸送黑色素給約 40 個角質細胞。慢性日曬皮膚黑色素細胞數目可達兩倍且活性增加;曬黑主要靠每細胞色素合成改變而非細胞數變動。表皮黑色素細胞可由毛囊黑色素細胞幹細胞 (melanocyte stem cells, McSCs)「儲庫」再生。
- 毛囊黑色素細胞:McSCs 駐留於隆突 (bulge)/次級毛胚,休止期靜止;生長期 (anagen) 經 Wnt 訊號活化、增殖、分化、產生色素;退化期 (catagen) 凋亡。McSC 隨年齡流失導致毛髮灰白 (hair graying),游離輻射等 DNA 損傷可加速。
- 肢端黑色素細胞:外分泌汗腺 (eccrine gland) 下部為肢端 McSC 微環境;肢端黑色素瘤可能源自此群體。
- 皮膚外黑色素細胞:黏膜、耳蝸、葡萄膜、軟腦膜、心臟;耳蝸血管紋 (stria vascularis) 缺陷解釋 Waardenburg 與 Tietz 的感音性耳聾。視網膜色素上皮源自神經外胚層、與黑色素細胞譜系無關。
黑色素細胞的形成、遷移與存活
- 源自高度遷移、多能 (multipotent) 神經嵴。傳統認為僅走背外側 (dorsolateral) 路徑;新證據顯示有第二波黑色素細胞源自許旺細胞前驅細胞 (Schwann cell precursors),沿腹內側 (ventromedial) 路徑遷移。
- 小鼠發育:黑色素母細胞 (melanoblasts) 自神經管分層後不久即被指定;MITF 約於胚胎第 10.5–11 日 (E10.5-11) 首次表現;經遷移待命區 (MSA) 後於 E12.5 沿背外側遷移;E14.5 表現酪胺酸酶、E16.5 顯示色素。神經嵴細胞保留發育可塑性 (developmental plasticity),此特性可能參與黑色素瘤發展。
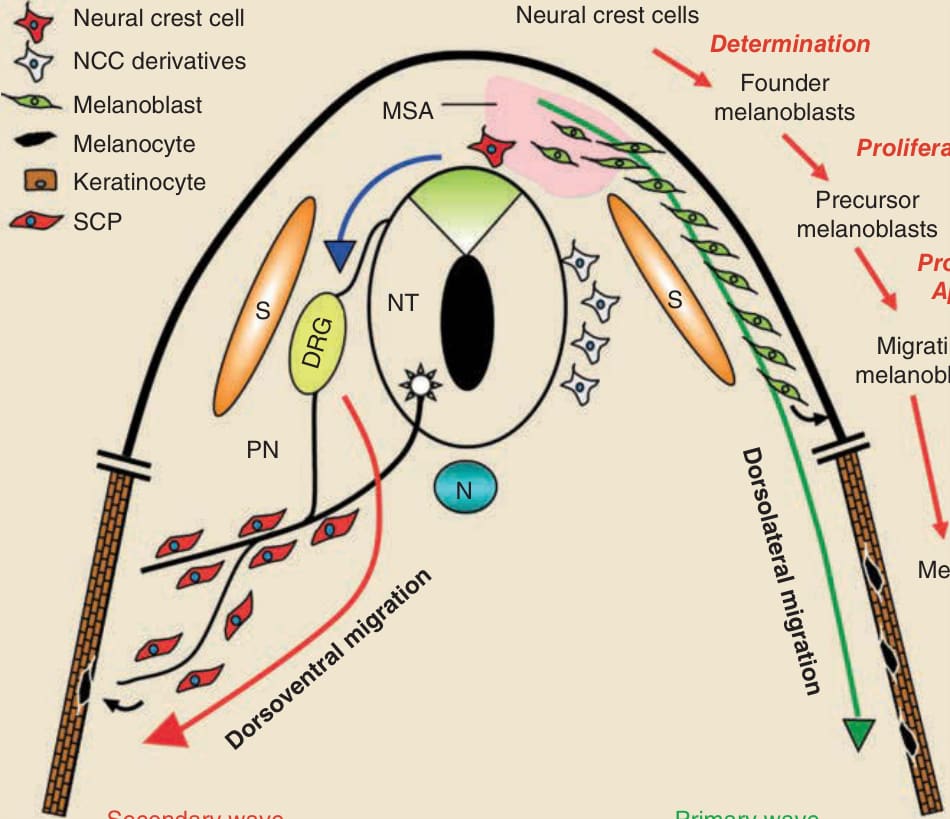
圖 20-3:發育期間黑色素細胞的指定與遷移(背外側初級波與許旺細胞前驅細胞衍生的腹側第二波)
- 調節因子:
- 轉錄因子:
- MITF(小眼畸形相關轉錄因子):黑色素細胞主控因子;受 SOX10、PAX3 正向調節;結合 E-box (CA[C/T]GTG) 驅動色素與黑色素體基因(酪胺酸酶、TYRP1、DCT、PMEL17、MLANA、Rab27a)、增殖基因 (CDK2)、存活基因 (BCL2)。為黑色素細胞存活、增殖、分化所必需;作為致癌基因,E318K 多型性(破壞 SUMO 化位點)增加黑色素瘤風險。
- PAX3、SOX10:神經嵴早期標記,正向調節 MITF;突變致 Waardenburg 症候群(並含非色素表型,如 SOX10 巨結腸症)。
- SOX2、FoxD3:抑制 MITF,偏好許旺細胞命運;其下調偏好黑色素細胞命運。
- 生長因子訊號:
- Ednrb(B 型內皮素受體):配體 Edn3 為發育關鍵;主要負責黑色素母細胞遷移。
- c-Kit / KITLG(幹細胞因子 SCF):c-Kit 異型合子突變致局部白斑;對成熟黑色素細胞存活與色素沉著重要;KITLG 多型性致 FPH/FPPH;活化突變/擴增驅動肢端雀斑樣、黏膜、惡性小痣黑色素瘤。
- Wnt/β-Catenin:典型 Wnt 經 β-連環蛋白活化 MITF,促進增殖與分化;亦增強黑色素瘤形成與轉移。
- Neuregulin/ErbB2/ErbB3:neuregulin 促許旺細胞命運、抑黑色素細胞命運。
- 運動/遷移:Rac1(Rho GTP 酶)調節肌動蛋白細胞骨架與遷移;活化突變見於 9% 日曬黑色素瘤。
- 轉錄因子:
黑色素與黑色素體
- 兩種黑色素皆源自前驅物酪胺酸 (tyrosine);酪胺酸酶將酪胺酸氧化為多巴醌 (dopaquinone),是調節真黑色素與褐黑色素生成的關鍵酶,需結合銅 (copper)(由 ATP7A 輸送)。
- 真黑色素(棕/黑)由 5,6-二羥基吲哚 (DHI) 與 DHICA 聚合而成;褐黑色素(黃/橙)由含硫半胱胺醯多巴 (cysteinyldopa) 氧化聚合而成,高度溶於鹼。
- 臨床意義:真黑色素屏蔽 UV 並清除活性氧物種 (ROS),保護對抗曬傷與皮膚癌(白化症與淺膚色者皮膚癌風險高)。褐黑色素屏蔽力差且可作為促氧化劑,UVA 與之反應產生自由基並造成「暗 CPD(dark cyclobutane pyrimidine dimer)」;在 BRAF(V600E) 小鼠模型中,Mc1r 功能障礙(紅髮表型)即使無 UV 也增加黑色素瘤風險,導入酪胺酸酶功能喪失(白化-紅髮)可消除此效應,顯示褐黑色素途徑可經 UV 依賴與非依賴機轉促成黑色素瘤。
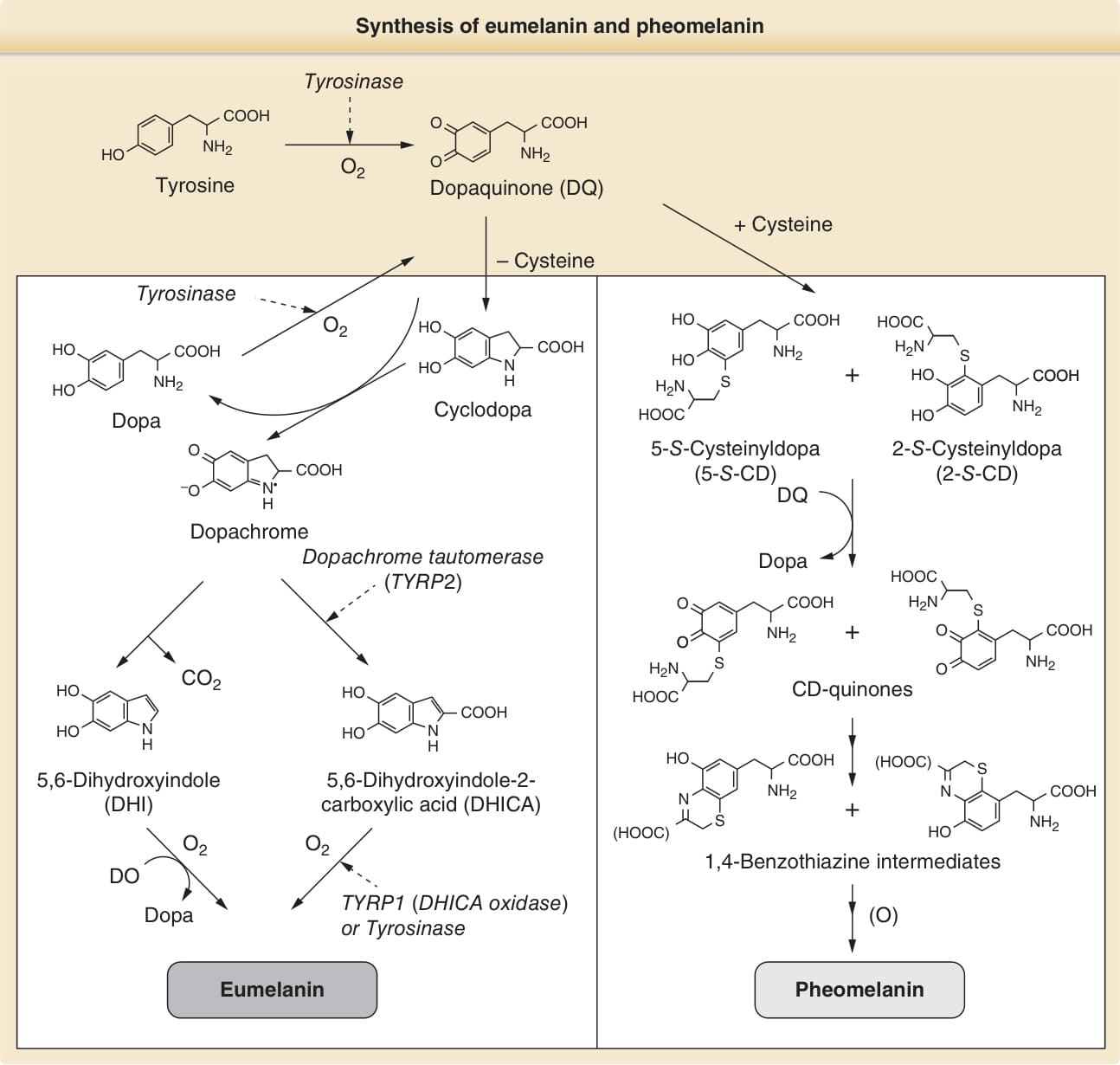
圖 20-5:真黑色素與褐黑色素的合成(共同前驅物 dopaquinone;TYRP1/TYRP2 參與真黑色素合成)
- 合成酶:TYRP1(DHICA 氧化酶)與 TYRP2/多巴色素互變異構酶 (DCT) 與酪胺酸酶同源,與酪胺酸酶形成複合體並穩定之;缺陷致眼皮膚白化症。DCT 偏好 DHICA 生成。
- 黑色素體生成與運輸:黑色素體源自內體途徑但需獨特分選途徑。PMEL(gp100/silver)構成纖維基質,於第 I/II 期被 BACE1 切割形成類澱粉纖維。黑色素生成酶(酪胺酸酶、TYRP1、DCT)經回收內體 (recycling endosome) 輸送。BLOC-1/2/3 與 AP-3 複合體(10 個基因突變致 Hermansky-Pudlak 症候群)負責貨物輸送;VAMP7 為 BLOC-1 依賴性 TYRP1 輸送所需的 SNARE,BLOC-3 負責 VAMP7 回收。
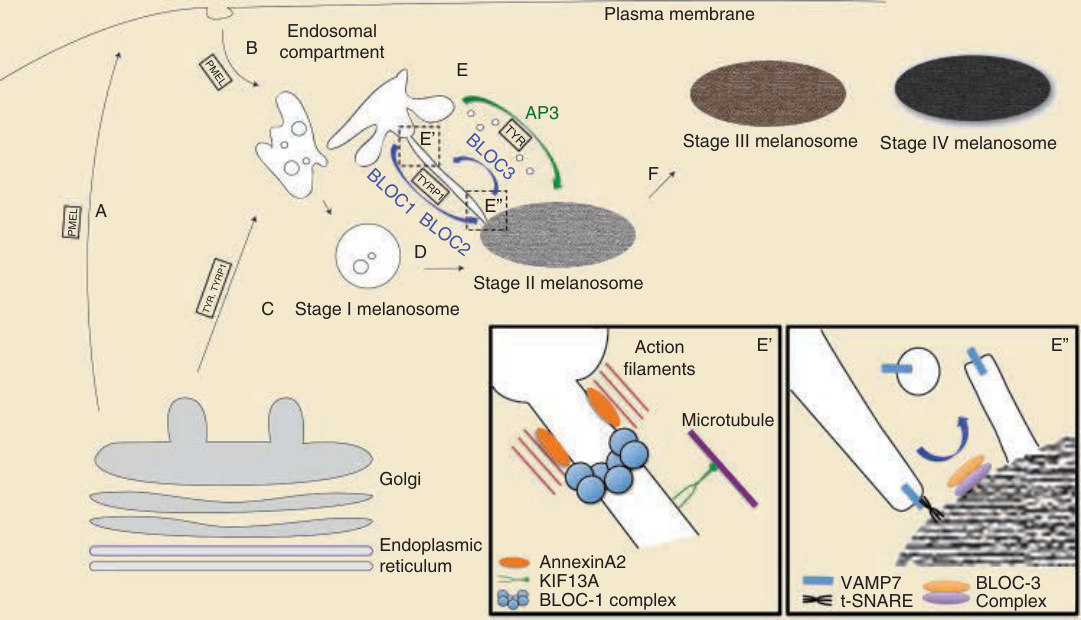
圖 20-6:黑色素體生成(內體途徑、PMEL 加工、BLOC/AP-3 介導的貨物輸送)
- 轉移至角質細胞:第 IV 期黑色素體經驅動蛋白 (kinesin) 順行運輸至樹突尖端;Rab27a–melanophilin (Mlph)–MyosinVa 複合體將黑色素體捕獲於肌動蛋白尖端(此複合體任一組成突變致 Griscelli 症候群)。轉移以脫落-吞噬 (shedding-phagocytosis) 模型證據最強。KGF 與 PAR2 活化增加角質細胞吞噬與攝取;PAR2 在深膚色者角質細胞中活性最高。
- 轉移後黑色素體經逆行運輸至核周形成「核罩 (nuclear capping)」保護 DNA。淺膚色者黑色素體小且成簇(膜包 4–8 個),深膚色者大、多、個別分散;深膚色者角質細胞降解黑色素體較慢(自噬受損)。
人類色素沉著變異的調節
- MSH-MC1R-MITF 途徑:MC1R 為黑色素細胞表現的 G 蛋白偶聯受體(7 個跨膜結構域),決定主要產生真黑色素或褐黑色素。失活變異致紅髮表型;活化突變致毛色加深。
- 配體:典型致效劑為 α-黑色素細胞刺激素 (α-MSH),源自前腦啡黑色素皮質素 (POMC) 切割(同時產生 β-腦內啡、ACTH);Agouti 蛋白 (ASIP) 為拮抗劑/反向致效劑,其變異與人類日曬敏感、雀斑、紅髮及皮膚癌相關。
- 訊號傳遞:MC1R 活化 → Gαs → 腺苷酸環化酶 → cAMP → PKA → 磷酸化 CREB → 上調 MITF → 黑色素生成。PDE4 為負回饋(抑制可刺激紅髮小鼠色素轉向棕/黑);PGC-1α/β 經 MITF 刺激黑色素生成。
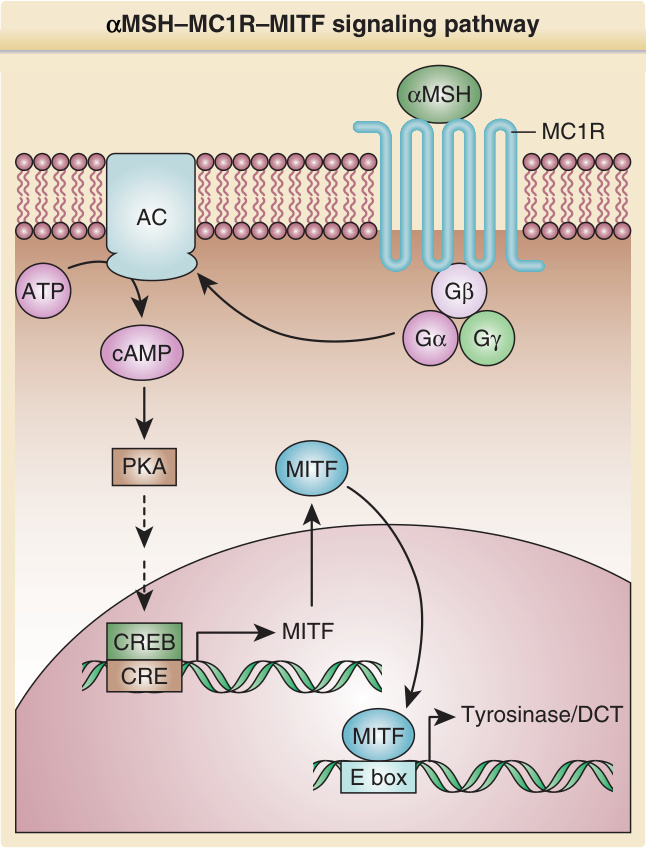
圖 20-10:αMSH–MC1R–MITF 訊號傳遞途徑(cAMP/PKA/CREB → MITF → 色素基因)
- UV 誘導曬黑:分立即曬黑(數分至數小時,UVA,現有黑色素氧化重分布)與延遲曬黑(UVB 為主,3 天至 1 週達峰,黑色素細胞數適度增加、樹突增加、黑色素化增強,保護作用較強)。角質細胞 UV → p53 依賴性上調 POMC → α-MSH → MC1R/MITF/黑色素生成。在紅髮、無法曬黑小鼠中局部施用毛喉素 (forskolin) 可繞過缺陷 MC1R、刺激下游 cAMP/PKA/MITF,增加色素並保護對抗 UV 致癌,顯示藥理逆轉紅髮表型的可能。UV 亦降低角質細胞 TGFβ1(MITF 的負調節因子)而增加色素。
- MC1R 與黑色素瘤生成:弱化型 (hypomorphic) MC1R 變異使黑色素瘤風險增加 2 至 4 倍;即使校正膚色仍增加風險,顯示除 UV 屏蔽減少外另有機轉(褐黑色素 ROS、核苷酸切除修復 NER 受損、UVB 介導 PTEN 降解增加)。
- 其他因子:族群遺傳學辨識出 SLC24A5(歐洲族群近普遍、致較淺膚色)、TPCN2(調節黑色素體 pH/大小)、IRF4(與 MITF 共同活化酪胺酸酶)、BNC2 (basonuclin 2) 等與膚色/髮色/眼色變異相關。
結論
- 黑色素細胞雖屬少數表皮細胞,卻對皮膚外觀與功能有重大影響;既組成型也回應環境合成色素。UV 保護主要來自真黑色素,褐黑色素及其合成途徑反而提升氧化壓力與致癌性。未來治療方向包括以 MSH 胜肽類似物或 MC1R 下游小分子,在淺膚色(MC1R 變異)個體中挽救真黑色素合成,以改善皮膚癌與光損傷預防。