毛囊生物學 (Biology of Hair Follicles) 精華筆記
重點一覽
- 毛髮僅見於哺乳動物,演化上原為隔熱與保護;在當代人類,主要角色是社交互動。禿髮 (alopecia) 與毛髮過多 (hirsutism、hypertrichosis) 造成顯著心理困擾。
- 毛囊發育取決於上皮細胞 (epithelial) 與間質細胞 (mesenchymal) 的交互作用;發育相關基因在毛囊週期 (hair follicle cycling) 中也扮演角色。
- 毛囊隆突 (bulge) 內有對毛囊持續再生至關重要的幹細胞 (stem cells)。
- 毛髮色素沉著取決於毛囊內的黑色素細胞幹細胞 (melanocyte stem cells) 與分化細胞。
胚胎學與發育機轉
- 毛囊發育分為八個連續階段。許多果蠅 (Drosophila) 發育基因的哺乳動物同源基因 (homologs) 也調控毛囊發育:Dpp/BMP、engrailed、Homeobox (hox)、hedgehog/patched、notch、wingless/wnt/catenin、branchless (Fgf)。
- 毛囊形成於子宮內由頭部開始向下延伸。最先形成的是胎毛 (lanugo hairs),無色素、柔軟細緻,通常於妊娠第 32 至 36 週脫落(約三分之一新生兒出生後仍保留數週)。
- 同源異形盒基因 (homeobox genes) 負責毛囊的非隨機對稱分布。Engrailed 缺乏的小鼠會在腳掌墊長出毛囊。
- 控制毛髮長度、捲曲度、分布的基因(來自狗的研究):FGF-5(長度)、Keratin 71(捲曲度)、R-spondin 2(分布)。亞洲人較粗毛髮與 Edar 活性增加相關。
毛囊型態發生的關鍵訊號
- WNT/β-catenin:最早期正向調控毛囊起始的途徑;β-catenin 為 WNT 下游介質,轉位入核與 LEF/TCF 形成複合體。β-catenin 活化建立「上皮勝任性 (epithelial competence)」。成年表皮人為活化 β-catenin 可使毛囊重新 (de novo) 發育;但持續活化會導致毛母質瘤 (pilomatricoma) 與毛囊瘤 (trichofolliculoma)。
- EDA/EDAR:促進早期毛囊發育。EDA 突變導致 X 染色體連鎖無汗性外胚層發育不全 (X-linked anhidrotic ectodermal dysplasia)(毛囊減少、牙齒與汗腺缺陷);EDAR 突變導致少汗性外胚層發育不全。EDAR 活性增加與較粗毛髮相關。
- BMP:抑制毛囊形成;其拮抗劑 Noggin 中和 BMP 可促進基板命運(部分透過上調 Lef-1)。Noggin 缺乏小鼠毛囊較少且發育延遲。
- Notch(配體 delta-1):促進並加速基板形成,同時抑制周圍細胞。
- SHH (Sonic hedgehog):受體為 Patched1 (PTC1);Shh 缺乏小鼠毛囊極度衰退、真皮乳頭發育不良。Patched 是基底細胞母斑症候群 (basal cell nevus syndrome) 缺陷基因。
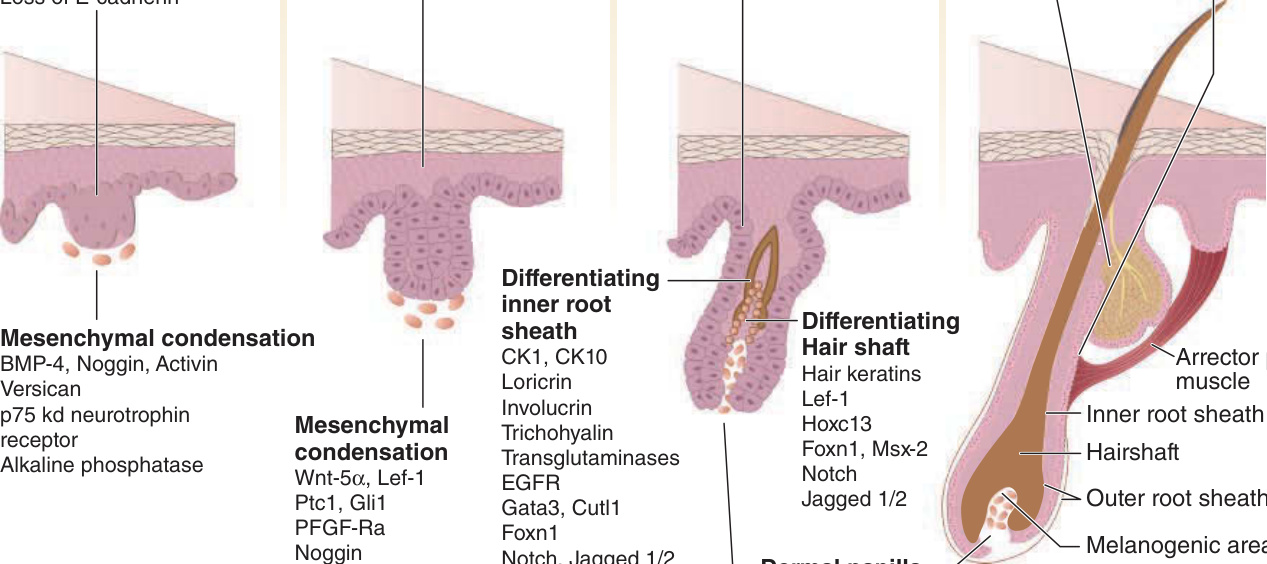
圖 7-1:毛囊形態發生的分子調控
毛囊發育階段
- 上皮基板 (epithelial placode)/初級毛胚芽:對應第 1 階段,約妊娠 10 週首次出現。基底表皮細胞向心性遷移與緊密化形成,表現胎盤鈣黏蛋白 (placental cadherin),喪失橋粒與上皮鈣黏蛋白。下方真皮細胞形成凝集 (condensation),發育為真皮乳頭 (dermal papilla)。
- 球狀栓/毛芽 (bulbous peg/hair bud):第 2 階段,毛胚芽延長為上皮細胞索。最深部形成毛母質 (matrix),產生毛幹與內毛根鞘。外毛根鞘形成兩個隆突:淺層發育為皮脂腺 (sebaceous gland),深層為上皮幹細胞所在;豎毛肌 (arrector pili) 附著於隆突區,收縮造成雞皮疙瘩。腋下、肛門生殖器、乳暈等部位有第三隆突發育為頂泌汗腺 (apocrine gland)。
- 成熟毛囊:至少八個細胞層形成。內毛根鞘分化由 EGFR 訊號與其配體 TGF-α 調節。毛幹分化受 Wnt、Bmp、Edar、FGF、Hedgehog、IGF、Notch 與多種轉錄因子(Foxn1、Lef1、Hoxc13 等)調控。人類頭皮毛囊主要叢集成三個一組。
解剖學
毛髮類型
- 終毛 (terminal):直徑 >60 µm,有中央髓質,可長 >100 cm,生長期 (anagen) 毛球位於皮下脂肪。
- 毳毛 (vellus):直徑 <30 µm,無髓質,<2 cm,毛球位於網狀真皮。
- 青春期下,男性生殖器、腋下、軀幹、鬍鬚的毳毛毛囊在性荷爾蒙影響下轉為終毛毛囊;頭皮終毛毛囊在雄性禿 (androgenetic alopecia) 過程轉為微小化毛囊。
- 毛髮彎曲度源自彎曲的毛囊;內毛根鞘形狀決定毛髮形狀。EGFR 途徑與 IGFBP5 突變導致小鼠捲髮。
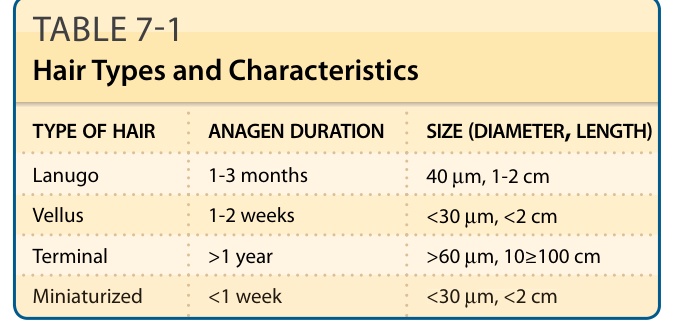
表 7-1:毛髮類型與特性
顯微解剖
- 上段毛囊(毛漏斗部 infundibulum+峽部 isthmus)為永久性;下段毛囊(球上部+球部)隨每個週期再生。
- 外毛根鞘 (outer root sheath):於毛漏斗部與表皮相連。峽部以毛鞘式 (trichilemmal) 角化、缺顆粒層(毛鞘性囊腫保留此特性)。隆突位於峽部基部。
- 內毛根鞘 (inner root sheath):由外到內為伴隨層 (companion layer)、亨利層 (Henle)、赫胥黎層 (Huxley)、角質層。亨利層最先角化;赫胥黎層於 Adamson 緣角化。由硬角蛋白組成,引導決定毛髮形狀。GATA-3 對其分化至關重要,缺乏者無法形成內毛根鞘。
- 毛幹 (hair shaft):源自毛球快速增殖的毛母質角質細胞(全身最高增殖率之一),形成髓質、皮質、角質層。分化依賴 Lef-1;BMP 受體 1a 型對毛母質分化為毛幹至關重要。皮質含黑色素,髓質角蛋白受雄性素 (androgens) 控制。
- 真皮乳頭 (dermal papilla):間質衍生核心,含纖維母細胞、膠原束、富黏多醣基質、神經纖維與單一微血管環。具強大誘導特性,體積與毛母質細胞數及毛幹大小相關。KGF 由生長期真皮乳頭產生(受體 FGFR2 於毛母質);但 KGF 基因剔除小鼠仍能形成正常毛囊(顯示可被冗餘因子取代)。
- 毛囊神經支配:有髓鞘感覺神經纖維環繞毛囊。神經末梢:自由神經末梢(疼痛)、披針狀末梢(加速度)、梅克爾細胞 (Merkel cells)(壓力)、毛-魯菲尼小體(張力)。
- 毛囊周圍鞘:內層基底膜(玻璃樣膜)+外層第 III 型膠原結締組織鞘。移植後可形成新真皮乳頭並誘導新毛囊,且無免疫排斥。
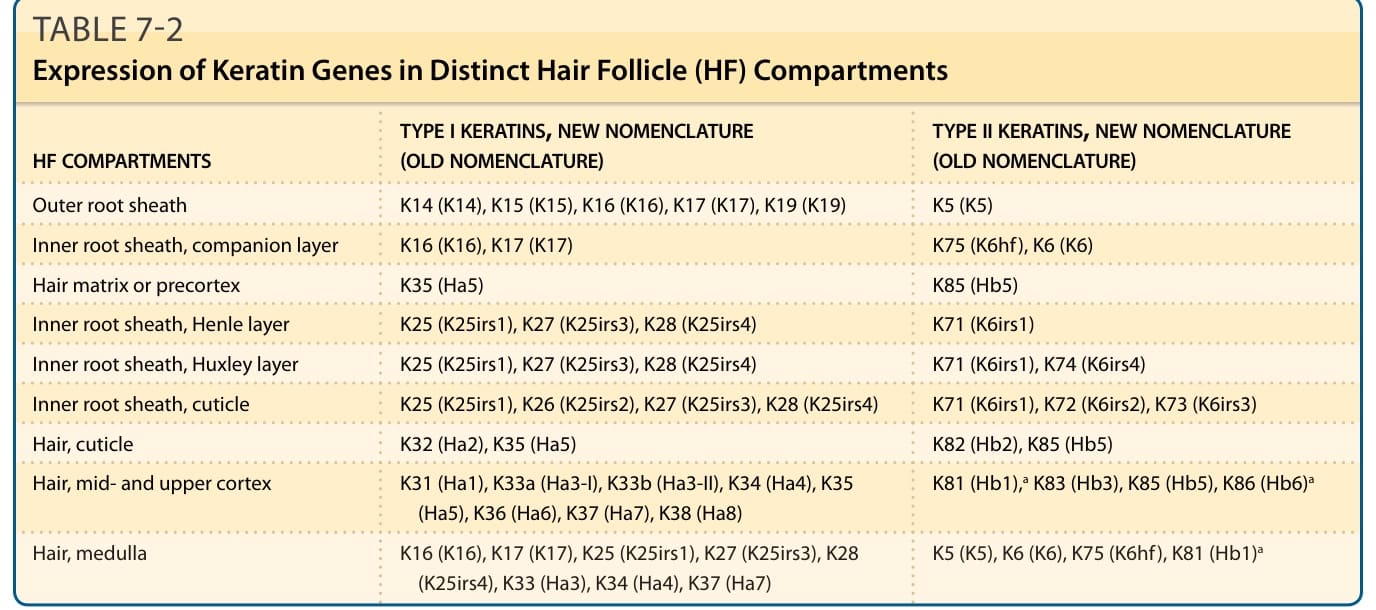
表 7-2:角蛋白基因在不同毛囊區室的表現
毛囊週期
- 三階段:生長 (anagen)、退化 (catagen)、休止 (telogen)。生長期長度決定毛髮最終長度。
- 生長期長度:頭皮 2 年至超過 8 年;腿部 5 至 7 個月;手臂 1.5 至 3.0 個月;睫毛 1 至 6 個月;手指 1 至 3 個月。
- 成年人類頭皮毛髮非同步生長,約 90% 至 93% 處於生長期。頭皮有 100,000 至 150,000 根毛髮,約 10,000 根處於休止期;每天僅掉落 50 至 100 根,故休止期為異質性狀態。
- 頭皮毛髮生長速率 0.37 至 0.44 mm/天,約 1 cm/月。
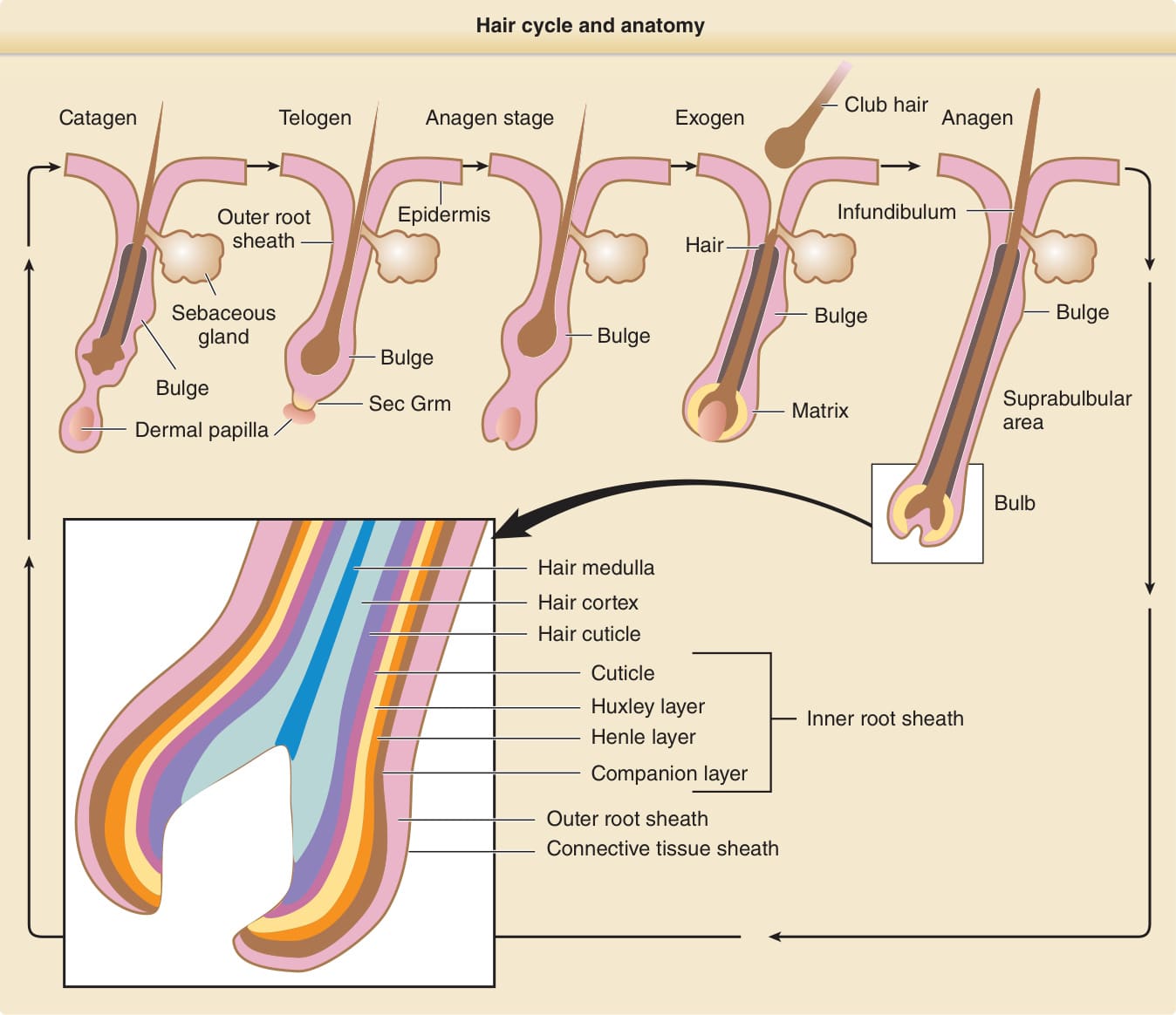
圖 7-2:毛髮週期與解剖
毛囊幹細胞
- 幹細胞永久位於隆突 (bulge)(非舊說的「次級胚芽」),譜系分析證實隆突細胞產生毛囊所有上皮層;隆突細胞消融導致毛囊破壞。隆突幹細胞喪失導致瘢痕性禿髮 (cicatricial alopecia)。
- 標記:cytokeratin 15、Lgr5(也標記隆突細胞)、Lgr6(上峽部,恆定與傷後遷移至表皮)。
- 全層傷後,隆突細胞後代遷移至傷口,約佔再生表皮細胞的 30%。
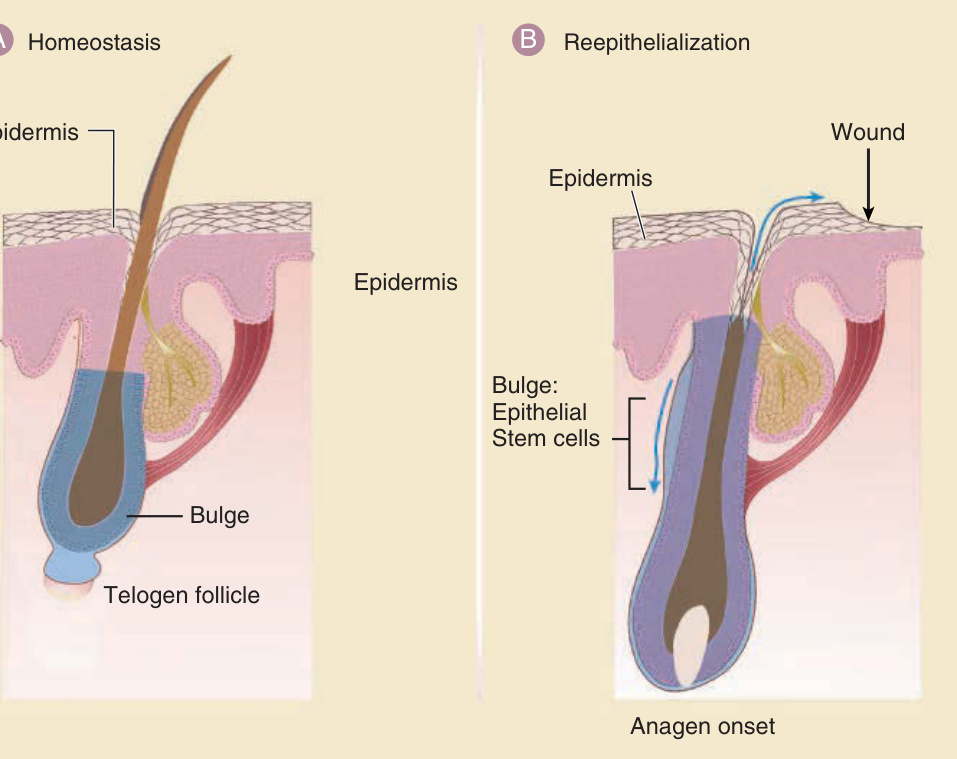
圖 7-3:恆定狀態與傷後表皮幹細胞與毛囊幹細胞之關係
生長期 (anagen)
- 重演胎兒期毛囊發生,分為七個階段(第 IV 期母質黑色素細胞重新活化)。
- 角質細胞遷移時出現蛋白酶與膠原酶,類似上皮傷口;Stat3 缺乏小鼠有傷口癒合缺陷且毛囊無法進入生長期。最後內皮增殖與血管新生標誌下段毛囊完全恢復。
退行期 (catagen)
- 母質細胞有絲分裂停止、協調性凋亡 (apoptosis)。色素生成先於增殖停止而停止(休止期棍狀毛近端無色素)。下段毛囊與真皮乳頭向上回縮。最大毛囊(頭皮)由 2 至 5 mm 縮短為休止期 0.25 至 0.5 mm。
- 真皮乳頭(受 Bcl-2 保護)凝集上移至隆突下方。真皮乳頭由皮下脂肪遷移回真皮對持續週期是必需的——丘疹性無毛症 (atrichia with papules) 即因 hairless 或維生素 D 受體基因突變致此遷移失敗(後者並伴佝僂病 rickets)。
- Fgf5:缺乏者毛髮比野生型長 50%(安哥拉表型 angora);於退行期前在外毛根鞘上調,可能觸發退行期。
- EGFR/TGF-α:對綿羊投予 EGF 誘導退行期;TGF-α 缺乏小鼠呈波浪狀毛髮 (waved hair);EGFR 下調使毛髮波浪且較長、生長期至退行期轉變延遲。
- 神經營養素與 TGF-β1:誘導過早退行期(BDNF 過度表現使毛髮縮短 15%)。
休止期與脫落期 (telogen and exogen)
- 退行期完成形成棍狀毛;約 1% 休止期毛髮每天脫落。脫落期 (exogen) 為主動階段。
- 立即休止期釋放 (immediate telogen release):可見於開始服用 minoxidil 等藥物後,棍狀毛被主動脫落。雄性禿病人脫落與再生長間的「滯後期」常持續數月。
毛髮色素沉著
- 毛髮著色源於黑色素合成與從毛球黑色素細胞運輸至毛幹角質細胞的協調程式,嚴格與生長期偶聯,退行期與休止期停止。
- 黑色素細胞發育:黑色素母細胞 (melanoblasts) 約妊娠 50 天可辨識,源自神經嵴 (neural crest) 與皮膚神經中的許旺細胞祖細胞。Pax3 與 Mitf 調節譜系承諾。Ednrb 與 c-kit 突變分別見於赫希施普龍病 (Hirschsprung disease) 與斑駁病 (piebaldism),導致無色素毛髮。SCF/c-kit 訊號為黑色素母細胞遷移與分化所必需。
- 黑色素細胞幹細胞 (MCSC):位於隆突,呈靜止狀態,表現 Trp2、Bcl-2、Pax3。TGF-β 與 Notch 訊號維持其存活。Bcl-2 剔除小鼠因 MCSC 耗竭而進行性白髮(可被 SCF 過度表現補償)。WNT 活化驅動 MCSC 分化為產色素黑色素細胞。
- 產色素黑色素細胞:位於毛球真皮乳頭上方,表現 tyrosinase、Trp1、Trp2(小鼠)、pMel17(人類)。
- 週期依賴變化:生長期早中期黑色素細胞最大增殖;Notch 防止過早分化。退行期色素生成驟停,部分黑色素細胞凋亡或落入真皮乳頭。
- 毛色分子調控:MC-1R 調節黑(eumelanin)與黃(pheomelanin)色素平衡。α-MSH 結合 MC-1R 升高 cAMP、活化 MITF 促進真黑色素酵素合成。Agouti signal protein (ASP) 競爭抑制 MC-1R 致褐黑色素。人類毛囊存在 POMC–MC-1R 途徑(α-MSH、ACTH、β-endorphin 促黑色素生成)。
- 白髮:濾泡黑色素細胞對老化敏感,過早喪失致白髮。白髮毛球黑色素細胞較少、外毛根鞘不活化黑色素細胞顯著減少。ATM-kinase 缺乏使 MCSC 對異位分化敏感,加速白髮。
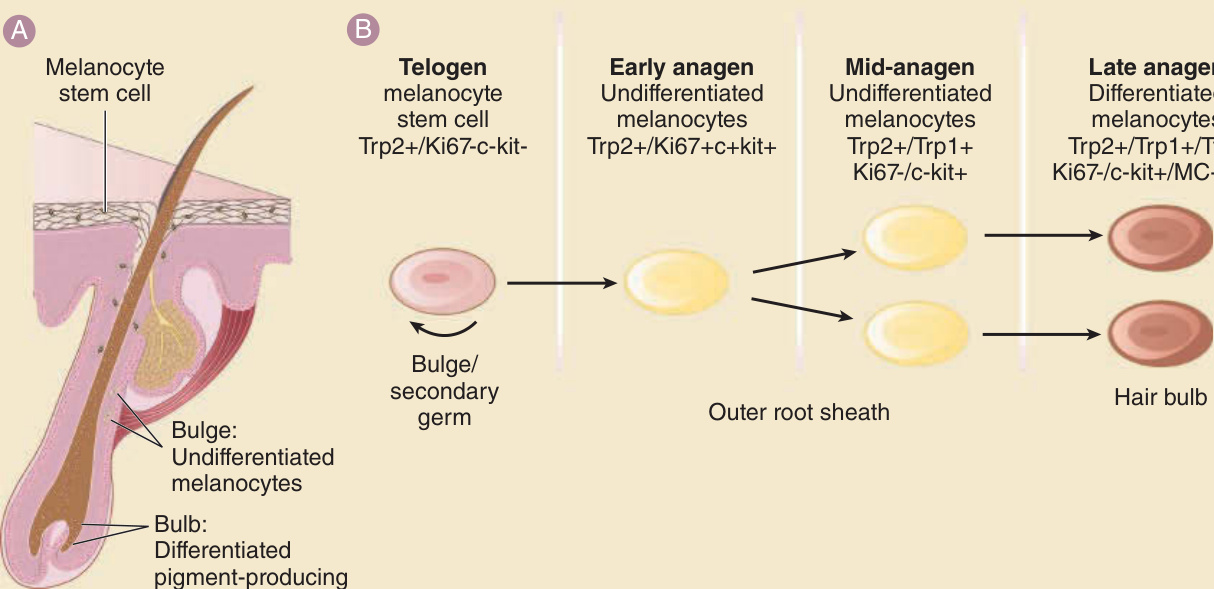
圖 7-5:毛髮週期依賴的濾泡色素單位重塑